
Locatie van lymfoïde weefsel. lymfoïde weefsel. Allergische reacties van het directe type
Allergie - overgevoeligheid organisme aan verschillende stoffen, geassocieerd met een verandering in zijn reactiviteit. Een kenmerk van allergische reacties is de verscheidenheid van hun klinische vormen en cursusvarianten.
Ze zijn ingedeeld in twee grote groepen: reacties van het directe type en reacties van het vertraagde type.
Allergische reacties van het directe type
ž Onmiddellijke reacties zijn onder meer anafylactische shock, Quincke's oedeem. Ze ontwikkelen zich letterlijk binnen een paar minuten nadat een specifiek antigeen (allergeen) het lichaam is binnengekomen. Quincke's oedeem (angioneurotisch oedeem) wordt gekenmerkt door zijn specifieke manifestatie in met name het gelaat.
Angio-oedeem (Quincke-oedeem)
Het treedt op als gevolg van de werking van voedselallergenen, verschillende medicijnen die oraal worden gebruikt, met plaatselijke toepassing. Gelokaliseerde ophoping van een grote hoeveelheid exsudaat in het bindweefsel, meestal in het gebied van de lippen, oogleden, slijmvlies van de tong en strottenhoofd. Oedeem verschijnt snel, heeft een elastische consistentie; weefsels in het oedeemgebied zijn gespannen; duurt enkele uren tot twee dagen en verdwijnt spoorloos en laat geen veranderingen achter. Angio-oedeem van het gezicht of alleen de lippen wordt vaak gezien als een geïsoleerde manifestatie van medicijnallergie. Het moet worden onderscheiden van: zwelling van de lippen met het Melkersson-Rosenthal-syndroom, Meige-trooedeem en andere macrocheilitis.
Quincke's oedeem, met een manifestatie op de bovenlip:
Met een manifestatie op de onderlip:
Vertraagde allergische reacties
Contact- en toxisch-allergische geneesmiddelenstomatitis
Ze zijn de meest voorkomende vorm van laesies van het mondslijmvlies bij allergieën. Ze kunnen optreden bij het gebruik van elk medicijn.
Klachten: branderig gevoel, jeuk, droge mond, pijn bij het eten. De algemene toestand van patiënten is in de regel niet gestoord.
Objectief: hyperemie en oedeem van het mondslijmvlies worden opgemerkt, op de laterale oppervlakken van de tong en wangen langs de lijn van het sluiten van de tanden, zijn de afdrukken van de tanden duidelijk zichtbaar. De tong is hyperemisch, helderrood. De papillen kunnen hypertrofisch of geatrofieerd zijn. Tegelijkertijd kan catarrale gingivitis optreden.
Differentiële diagnostiek: vergelijkbare veranderingen in de pathologie van het maagdarmkanaal, hypo- en avitaminose C, B1, B6, B12, endocriene stoornissen, diabetes mellitus, CCC-pathologie, schimmelinfecties.
Geneesmiddelstomatitis, met lokalisatie op de onderlip:

Medische catarrale gingivostomatitis, met lokalisatie op de bovenlip:

Ulceratieve laesies van het mondslijmvlies
♠ž Optreden tegen de achtergrond van oedeem en hyperemie in de lippen, wangen, laterale oppervlakken van de tong, hard gehemelte.
♠ž Er zijn erosies van verschillende groottes, pijnlijk, bedekt met fibrineuze plaque.
♠ž Erosies kunnen met elkaar versmelten en een uniform erosief oppervlak vormen.
♠ž De tong is bedekt met een coating, oedemateus. Gingivale interdentale papillen zijn hyperemisch, oedemateus en bloeden gemakkelijk bij aanraking.
♠ž Submandibulaire lymfeklieren zijn vergroot, pijnlijk. De algemene toestand is verstoord: koorts, malaise, gebrek aan eetlust.
♠ž Differentiële diagnose: het is noodzakelijk om te differentiëren van herpetische stomatitis, afteuze stomatitis, pemphigus, erythema multiforme.
Medische erosieve stomatitis:

Ulceratieve-necrotische laesies van het mondslijmvlies
♠ž Het proces kan worden gelokaliseerd op het harde gehemelte, tong, wangen.
♠ž Het kan diffuus zijn, waarbij niet alleen het mondslijmvlies betrokken is, maar ook de palatinale amandelen, de achterste faryngeale wand of zelfs het gehele maagdarmkanaal.
♠ž Zweren zijn bedekt met witgrijs necrotisch verval.
♠ž Patiënten klagen over hevige pijn in de mond, moeite met het openen van de mond, pijn bij het slikken, koorts.
♠ž Differentiële diagnose: ulceratieve necrotische stomatitis van Vincent, traumatische en trofische ulcera, specifieke laesies bij syfilis, tuberculose, evenals ulceratieve laesies bij bloedziekten.
Geneesmiddel ulceratieve necrotische stomatitis met lokalisatie op het onderoppervlak van de tong:

Specifieke allergische manifestaties op het slijmvlies, bij het nemen van bepaalde medicijnen
♠žž Vaak verschijnen als gevolg van het innemen van een medicinale stof blaren of blaren op het slijmvlies van de mondholte, na opening waarvan zich meestal erosies vormen. Dergelijke huiduitslag wordt voornamelijk waargenomen na het nemen van steptomycine. Soortgelijke elementen op de tong, lippen kunnen verschijnen na inname van sulfonamiden, oletetrina.
♠žž Veranderingen in de mondholte als gevolg van het nemen van tetracycline-antibiotica worden gekenmerkt door de ontwikkeling van atrofische of hypertrofische glossitis
♠žž Laesies van de mondholte gaan vaak gepaard met schimmelstomatitis.
Veranderingen in de mondholte als gevolg van het nemen van sulfonamiden in de vorm van oedeem en hyperemie van de bovenlip en een gebied van necrose op het slijmvlies van de tong:


De reactie van het slijmvlies op oletethrine in de vorm van erosies op de laterale oppervlakken van de tong:


De reactie van het mondslijmvlies op antibiotica in de vorm van papillaire hypertrofie, erosies in de tong en atrofie van de papillen, na inname van tetracycline (tetracycline tong):


Allergische purpura of Schönlein-Genuch-syndroom
♠ž Aseptische ontsteking van kleine bloedvaten door het schadelijke effect van immuuncomplexen.
♠ž Gemanifesteerd door bloedingen, verminderde intravasculaire bloedstolling en microcirculatiestoornissen.
♠ž Het wordt gekenmerkt door hemorragische uitslag op het tandvlees, de wangen. taal, lucht. Petechiën en hemorragische plekken met een diameter van 3-5 mm tot 1 cm steken niet uit boven het niveau van het slijmvlies en verdwijnen niet wanneer er met glas wordt gedrukt.
♠ž De algemene toestand van de patiënten is verstoord, zwakte, zorgen over malaise.
♠ž Differentiële diagnostiek: ziekte van Wergolf, homofilie, beriberi C.
Shenlein-Genyuha-syndroom:

Diagnose van contact- en toxisch-allergische geneesmiddelenstomatitis
♠ž Allergische geschiedenis.
♠ž Kenmerken van de klinische cursus.
♠ž Specifieke allergologische, huidallergische testen.
♠ž Hemogram (eosinofilie, leukocytose, lymfopenie)
♠ž Immunologische reacties.
Behandeling van contact- en toxisch-allergische geneesmiddelenstomatitis
♠ž Etiotrope behandeling - isolatie van het lichaam tegen de invloed van het vermeende antigeen.
♠ž Pathogenetische behandeling - remming van de proliferatie van lymfocyten en de biosynthese van antilichamen; remming van de antigeen-antilichaamverbinding; specifieke desensibilisatie; BAS-inactivering.
♠ž Symptomatische behandeling - invloed op secundaire manifestaties en complicaties (correctie van functiestoornissen in organen en systemen)
♠ž Specifieke hyposensibiliserende therapie wordt uitgevoerd volgens speciale schema's na een grondig allergologisch onderzoek en bepaling van de staat van sensibilisatie van de patiënt voor een specifiek allergeen.
♠ž Niet-specifieke hyposensibiliserende therapie omvat: calciumpreparaten, histoglobuline, antihistaminica (Peritol, Tavegil), evenals ascorbinezuur en ascorutine.
♠ž In ernstige gevallen worden corticosteroïden voorgeschreven.
♠ž Lokale behandeling wordt uitgevoerd volgens het principe van therapie voor catarrale stomatitis of erosief-necrotische laesies van de slijmvliezen: antiseptica met anesthetica, antihistaminica en corticosteroïden, ontstekingsremmende geneesmiddelen en proteïnaseremmers.
♠ž Proteolytische enzymen zijn geïndiceerd voor necrotische laesies;
♠ž Voor restauratie - keratoplastische preparaten.
Behcet-syndroom
♠ž Dento-oftalmogenitaal syndroom.
♠ž Etiologie: infectieuze allergie, auto-agressie, genetische conditionering.
♠ž Begint meestal met malaise, die gepaard kan gaan met koorts en spierpijn.
♠ž Aften verschijnen op de slijmvliezen en slijmvliezen van de uitwendige geslachtsorganen. Er zijn veel achterwanden, ze zijn omgeven door een inflammatoire rand van felrode kleur, hebben een diameter tot 10 mm. Het oppervlak van de aften is dicht opgevuld met geelwitte fibrineuze plaque.
♠ž Ze genezen zonder litteken.
♠ž Oogbeschadiging komt voor bij bijna 100% van de patiënten, gemanifesteerd door ernstige bilaterale iridocyclitis met vertroebeling van het glaslichaam, wat leidt tot de geleidelijke vorming van synechia, overgroei van de pupil.
♠ž In sommige gevallen verschijnt er uitslag op de huid van het lichaam en de ledematen in de vorm van erythema nodosum.
♠ž De meest ernstige complicatie is zenuwstelsel, die verloopt volgens het type meningo-encefalitis.
♠ž Andere symptomen van het syndroom van Behcet: de meest voorkomende zijn terugkerende epididymitis, gastro-intestinale laesies, diepe zweren die vatbaar zijn voor perforatie en bloeding, vasculitis.
Behandeling van het syndroom van Behcet
Er zijn momenteel geen algemeen aanvaarde behandelingen. Corticosteroïden hebben geen significant effect op het verloop van de ziekte, hoewel ze de manifestatie van sommige klinische symptomen kunnen verminderen. In sommige gevallen worden colchicine en levamisol gebruikt - wat alleen effectief is in verband met de mucocutane manifestaties van het syndroom. antibiotica voorschrijven een breed scala acties, plasmatransfusie, gamaglobuline.
Behcet-syndroom:

Erythema multiforme exsudatief
♠ž Een allergische ziekte met een acuut cyclisch verloop, vatbaar voor recidieven, gemanifesteerd door polymorfisme van huiduitslag en mondslijmvlies.
♠ž Het ontwikkelt zich voornamelijk na het nemen van medicijnen (sulfonamiden, ontstekingsremmende medicijnen, antibiotica) of onder invloed van huishoudelijke allergenen.
♠ž Gemanifesteerd door verschillende morfologische elementen: vlekken, papels, blaren, blaasjes en blaren.
♠ž De huid, het slijmvlies van de mondholte kunnen afzonderlijk worden aangetast, maar hun gecombineerde laesie komt ook voor.
♠ž Besmettelijke-allergische vorm van MEE - begint als een acute infectie. Maculopapulaire uitslag verschijnt op de huid, lippen, oedemateus en hyperemische mucosa. In de eerste stadia verschijnen blaren en blaasjes, die worden verdreven door sereus of sereus hemorragisch exsudaat. Elementen kunnen binnen 2-3 dagen worden waargenomen. De blaren scheuren en worden leeg, en in hun plaats worden talrijke erosies gevormd, bedekt met een geelgrijze fibrineuze coating (brandeffect).
♠ž Toxisch-allergische vorm van MEE - treedt op als een verhoogde gevoeligheid voor medicijnen wanneer ze worden ingenomen of ermee in contact komen. De frequentie van recidieven hangt af van contact met het allergeen. Bij deze vorm van de MEE is de ER een verplichte plaats voor het neerslaan van de elementen van de laesie. Huiduitslag is volledig identiek aan de vorige vorm, maar komt vaker voor en de fixatie van het proces is hier kenmerkend. Complicaties van deze vorm zijn conjunctivitis en keratitis.
♠ž Bij het diagnosticeren van MEE is het, naast de anamnese en klinische onderzoeksmethoden, noodzakelijk om een bloedtest uit te voeren, een cytologisch onderzoek uit te voeren van het materiaal uit de getroffen gebieden.
♠ž Differentiële diagnose: herpetische stomatitis, pemphigus, ziekte van Duhring, secundaire syfilis.
MEE. Erosies en korsten op de rode rand van de lippen en de huid van het gezicht:

MEE. Bubbels op het tandvlees en de slijmvliezen van de onderlip:

MEE. Erosies op het slijmvlies van de lippen, bedekt met fibrineuze plaque:

MEE. Erosies bedekt met een fibrineuze film op de lippen:


MEE. Uitgebreide erosies bedekt met een fibrineuze film op het onderoppervlak van de tong:

Kokarden:

Behandeling van exsudatief erythema multiforme
♠ Zorgt voor de identificatie en eliminatie van de sensibilisatiefactor.
♠ Voor de behandeling van een infectieus-allergische vorm wordt specifieke desensibilisatie uitgevoerd met microbiële allergenen.
♠ Het ernstige verloop van de ziekte is een directe indicatie voor de benoeming van corticosteroïden. loop van lysozym.
♠ Lokale behandeling wordt uitgevoerd, volgens de principes van therapie voor ulceratieve necrotische processen van mondslijmvlies - irrigatie met antiseptische oplossingen, oplossingen die de immunobiologische weerstand verhogen, geneesmiddelen die necrotische weefsels en fibrineuze plaque afbreken.
♠ Een kenmerk van de behandeling van MEE is het gebruik van geneesmiddelen die een lokaal anti-allergisch effect hebben (difenhydramine, thymaline) - in de vorm van toepassingen of een aerosol.
Stevens-Johnson-syndroom
♠ Ectodermose met lokalisatie nabij de fysiologische openingen.
♠ De ziekte is een falen van een superernstige vorm van exsudatief erythema multiforme, die optreedt met een significante verslechtering van de algemene toestand van de patiënten.
♠ Ontwikkelt zich als een medische laesie. Tijdens het ontwikkelingsproces kan het veranderen in het syndroom van Lyell. Het kan worden veroorzaakt door niet-steroïde anti-inflammatoire geneesmiddelen.
♠ De belangrijkste veranderingen vinden plaats in het integumentaire epitheel. Ze manifesteren zich door spongiose, ballondystrofie, in de papillaire laag van de lamina propria - het fenomeen van oedeem en infiltratie.
♠ Klinisch: de ziekte begint vaak met hoge temperatuur lichaam, vergezeld van vesiculaire en erosieve elementen van de laesie, ernstige schade aan de ogen met het verschijnen van bellen en erosies op het bindvlies.
♠ Een constant symptoom van het syndroom is een gegeneraliseerde laesie van het mondslijmvlies, vergezeld van het verschijnen van wijdverspreide erosies bedekt met witte vliezige plaque.
♠ Met een gegeneraliseerde laesie ontwikkelt zich vulvoaginitis.
♠ Huiduitslag wordt gekenmerkt door polymorfisme.
♠ Papels op de huid verzonken vaak in het midden, wat doet denken aan "kokardes"
♠ Op de rode rand van de lippen, tong, zacht en hard gehemelte vormen zich blaren met sereus-hemorragisch exsudaat, na lediging die uitgebreide pijnlijke erosies en foci verschijnen, bedekt met massieve purulent-hemorragische korsten.
♠ Mogelijke ontwikkeling van longontsteking, encefalomyelitis met fatale afloop.


Een allergie die zich manifesteert op het mondslijmvlies kan een zeer ernstig probleem worden. Over wat deze ziekten veroorzaakt en hoe ermee om te gaan, zullen we in dit artikel vertellen.
Waarom kan er een allergie zijn in de mondholte en eromheen?
Allergie op de lippenZoals u weet, ontstaat elke allergie doordat het menselijk immuunsysteem bepaalde stoffen niet goed kan waarnemen. Het geeft een signaal aan het hele lichaam om van het allergeen af te komen. Dientengevolge beginnen mestcellen een speciaal eiwit histamine te produceren, dat in het bloed door het lichaam wordt verdeeld en ontstekingsprocessen activeert. Dit is hoe allergieën beginnen.
De manier waarop het allergeen het lichaam binnenkomt, hangt af van wat de symptomen en manifestaties van een allergische reactie zullen zijn. Stuifmeel tast de luchtwegen en de slijmvliezen van neus en ogen aan. De chemicaliën kunnen de huid beschadigen. Het lichaam reageert volledig op dierenhaar. Voedselallergenen veroorzaken een reactie van het maagdarmkanaal en de huid. Hieruit kunnen we concluderen dat allergieën in de mond worden veroorzaakt door die stoffen die in direct contact staan met het mondslijmvlies, evenals de huid rond de lippen.
Zeer gemeenschappelijke oorzaak het optreden van de reactie is het nemen van medicijnen. Dit kunnen antibiotica, pijnstillers, jodiumpreparaten zijn. Daarnaast kan een reactie bij een volwassene worden veroorzaakt door tandheelkundige implantaten of prothesen. Bij een kind worden allergieën vaak veroorzaakt door voedingsmiddelen van de eerste aanvullende voedingsmiddelen, terwijl de reactie zich ook in de mond en rond de lippen kan ontwikkelen. Bovendien veroorzaakt de opkomst van problemen het wijdverbreide gebruik bij de productie van kleurstoffen, conserveermiddelen en andere chemicaliën die aan eindproducten worden toegevoegd en die ook worden gebruikt om groenten en fruit te verbouwen. Allergieën voor hen komen heel vaak voor.
Soorten allergische aandoeningen van het mondslijmvlies
Allergische aard heeft de volgende ziekten, die vrij vaak voorkomen:

Ziekte pathogenese
 Allergie in de mond: op de voorlip van binnenuit
Allergie in de mond: op de voorlip van binnenuit Er zijn verschillende classificaties van allergenen. Ze vormen allemaal twee grote groepen: exogeen en endogeen. Exogeen komt het lichaam van buitenaf binnen, terwijl ze besmettelijk en niet-infectieus van aard kunnen zijn. De tweede groep wordt gevormd door die allergenen die het lichaam zelf produceert als gevolg van schadelijke factoren. De methode van de behandeling hangt af van de aard van de ziekte.
Wanneer een allergeen binnendringt of een factor het lichaam beïnvloedt, kunnen verschillende soorten reacties optreden:
Anafylactisch (atopisch type)
cytotoxisch
immuuncomplex
Vertraagd
Het type ontwikkeling van een allergische reactie hangt af van de toestand van het lichaam, evenals de aard van de allergenen.
Allergiesymptomen in de mond en rond de lippen
Allergische aandoeningen van het mondslijmvlies en de huid rond de lippen kunnen symptomen hebben die niet lijken op typische allergieën. Meestal gaan ze gepaard met het verschijnen van kleine zweren, laesies van het slijmvlies op de wangen, lippen, zwelling van de tong. Bovendien kunnen de lippen vervellen en opzwellen, er vormen zich pijnlijke zweren en de huid rond de mondholte wordt aangetast.
 Allergische conjunctivitis kan optreden
Allergische conjunctivitis kan optreden Tegelijkertijd worden andere symptomen van een allergische reactie waargenomen, die het meest voorkomen:
allergische rhinitis
allergische conjunctivitis
Atopische dermatitis, eczeem, urticaria
Vernauwing van het lumen van de bronchiën, hoesten, kortademigheid
Quincke's oedeem
Anafylactische shock
Het is belangrijk om allergieën op tijd te herkennen, niet om zelf te proberen de ziekte te behandelen, omdat dit kan leiden tot een verergering van de aandoening. Als u tekenen van een allergie bij uw kind opmerkt, neem dan contact op met een allergoloog om het probleem effectief en zonder gevolgen op te lossen.
Diagnostiek
Diagnose is een belangrijke stap in de behandeling van allergieën, omdat het noodzakelijk is om te begrijpen op welke stof het lichaam zo heftig reageert. Zorg ervoor dat u uw arts vertelt wat u de afgelopen dagen heeft gegeten, welke medicijnen u gebruikt, welke chemicaliën mogelijk in contact zijn gekomen met het mondslijmvlies. Deze informatie is erg belangrijk.
Allergologen gebruiken verschillende methoden voor het diagnosticeren van ziekten. Ze zijn gebaseerd op hetzelfde mechanisme waarmee elke allergie zich ontwikkelt: het allergeen moet in de bloedbaan komen, waarna de resultaten worden geregistreerd. In laboratoriumomstandigheden is eenvoudig vast te stellen welke stof een verhoging van histamine in het bloed veroorzaakt en welke op geen enkele manier invloed heeft op de samenstelling van het bloed.
 U moet naar een kinderallergist
U moet naar een kinderallergist Na het eerste onderzoek kiest de allergoloog de diagnostische methode die bij u past. Dit kunnen prikkelende tests zijn, huidtesten, een bloedtest voor immunoglobulinespiegels. Het heeft geen zin voor kinderen onder de 3 jaar om tests te doen, omdat hun immuunsysteem niet voldoende is gevormd. Daarom is het gemakkelijker om de voeding van het kind aan te passen, evenals zijn omgeving, zodat er geen irriterende stoffen zijn. Dan zal de baby gezond opgroeien en geen allergiesymptomen voelen.
Behandelmethoden
Om de behandeling effectief te laten zijn, is het noodzakelijk om contact met het allergeen uit te sluiten. Daarom is de diagnose zo belangrijk. Zodra de arts begrijpt waardoor een dergelijke reactie van het immuunsysteem wordt veroorzaakt, kan een behandelplan worden opgesteld. Dit geldt vooral wanneer allergieën worden veroorzaakt door medicijnen. Daarom kunt u een kind niet behandelen met die medicijnen die de arts niet heeft voorgeschreven!
Therapie bestaat voornamelijk uit antihistaminica, die de samenstelling van het bloed normaliseren en het van histamine bevrijden. Dit leidt tot de verlichting van alle allergiesymptomen die tijdens de reactie optreden. Als het geval zeer ernstig is, is het noodzakelijk om corticosteroïdhormonen voor te schrijven. Bovendien is het raadzaam om sorberende preparaten te nemen, zodat het allergeen snel uit het lichaam wordt verwijderd. Dit zal uw herstel versnellen.
 Allergiecrème aanbrengen (op recept)
Allergiecrème aanbrengen (op recept) Om het ongemak in de mond te verminderen, kunt u speciale zalven, gels en antimicrobiële middelen, pijnstillers en antiseptica gebruiken om het mondslijmvlies te behandelen. Het is belangrijk om het proces niet te starten, zodat de ziekte niet chronisch wordt. Het is belangrijk om de huid rond de lippen te hydrateren om het genezingsproces te versnellen.
Er wordt een speciale behandeling voor het kind gekozen, omdat baby's niet veel medicijnen kunnen nemen, dus het belangrijkste is om een stabiele toestand te behouden en verslechtering te voorkomen. Als je merkt dat het gezicht van de baby begint te zwellen, bel dan onmiddellijk een ambulance! Dus Quincke's oedeem kan beginnen.
Preventie en voorzorgsmaatregelen
Om uzelf en uw kind te beschermen tegen allergieën, die zich niet alleen in de mond manifesteren, maar ook totaal andere symptomen kunnen hebben, is het belangrijk om de volgende voorzorgsmaatregelen in acht te nemen:
Kies voor een kind alleen die producten die geen kunstmatige toevoegingen bevatten. Het is het beste om groenten en fruit uit je eigen tuin te gebruiken voor aanvullende voedingsmiddelen. Speelgoed moet altijd schoon zijn, gebruik alleen hypoallergene waspoeders. Gebruik geen geparfumeerde cosmetica, vooral niet op het gezicht en de handen.
Neem geen medicijnen die u nog nooit heeft ingenomen zonder een arts te raadplegen. Geef uzelf of uw kind nooit zelfmedicatie. Het kan gevaarlijk zijn.
Tijdens de bloeiperiode goed je handen wassen en ook na elke wandeling je gezicht afspoelen. Hetzelfde geldt voor contact met dieren.
Als u merkt dat uw kind roodheid heeft op het gezicht, rond de mond, op het lichaam, analyseer dan welk voedsel de baby gedurende de dag heeft gegeten. Meld deze informatie aan de kinderarts om erachter te komen wat een dergelijke reactie kan hebben veroorzaakt.
Rijst. 17. Lokalisatie van plaatsen van lymfocytopoëse in het menselijk lichaam.
DE LYMFE KNOPEN ( nodi lympnatici) - een boonvormige verdikking langs de lymfevaten, waar antigeenafhankelijke reproductie van B- en T-lymfocyten plaatsvindt, hun verwerving van immuuncompetentie, evenals de zuivering van lymfe van vreemde deeltjes. De totale massa van lymfeklieren is 1% van het lichaamsgewicht, dat wil zeggen ongeveer 700 g. Lymfeklieren vormen meer dan 50 groepen. Volgens topografie zijn ze verdeeld in knooppunten van het lichaam (somatisch), ingewanden (visceraal) en gemengd, waarbij ze lymfe verzamelen van zowel de ingewanden als andere organen. De grootte van de lymfeklieren ligt in het bereik van 5-10 mm.
De lymfeklier is bedekt met een bindweefselcapsule, van waaruit bindweefselpartities zich uitstrekken tot in de orgel - trabeculae. In de capsule van sommige lymfeklieren werden gladde myocyten gevonden, die deelnemen aan de vorming van het bewegingsapparaat van de knoop.
Rijst. achttien. Diagram van de structuur van de lymfeklier. De rechterkant van de figuur toont de vascularisatie van de structurele elementen van het orgel.
Het parenchym van de knoop wordt gevormd door B- en T-lymfocyten, waarvan het skelet het reticulaire weefsel vormt. Er zijn corticale en medulla van de lymfeklier. De corticale substantie wordt gevormd door lymfatische follikels (knobbeltjes) die zich onder de capsule bevinden - bolvormige clusters van B-lymfocyten met een diameter van 0,5-1 mm. Naast B-lymfocyten omvat de samenstelling van de lymfeklierfollikels zowel typische macrofagen als een speciale variëteit van zichzelf, dendritische cellen genoemd. Uitwendig is de follikel bedekt met reticuloendotheliocyten - cellen die de morfologie van reticulaire cellen combineren met de functie van het endotheel, aangezien ze de sinussen van de lymfeklieren bekleden. Onder de reticuloendotheliocyten bevinden zich een aanzienlijk aantal vaste macrofagen, de zogenaamde kustcellen. Elke follikel bevat een licht (reactief of germinaal) centrum, waar lymfocyten zich vermenigvuldigen en waar B-lymfoblasten voornamelijk gelokaliseerd zijn, en een donkere perifere zone, waarin kleine en middelgrote lymfocyten compact zijn gelokaliseerd. Een toename van het aantal en de grootte van de reactieve centra van de follikels van de lymfeklieren duidt op antigene stimulatie van het lichaam.
Rijst. negentien. Lichtmicrofoto van een fragment van een lymfeknoop, x 200. Gekleurd met hematoxyline-eosine.
De medulla van de lymfeklier wordt gevormd door medullaire koorden - lintvormige clusters van B-lymfocyten, plasmacellen en macrofagen, langwerpig in de richting van de poort van de knoop naar de follikels. Uiterlijk zijn de hersenkoorden, evenals de follikels van de corticale substantie, bedekt met reticuloendotheliocyten. Tussen de hersenkoorden en follikels, respectievelijk, tussen de medulla en de cortex van de lymfeklier, is er een diffuse ophoping van T-lymfocyten, die de paracorticale zone wordt genoemd. Macrofagen in de paracorticale zone worden vertegenwoordigd door een verscheidenheid aan zogenaamde interdigiterende cellen die met elkaar in contact komen via vingervormige processen en stoffen produceren die de proliferatie van T-lymfocyten stimuleren. De corticale en medulla zijn dus bursa-afhankelijk en de paracorticale laag is de thymus-afhankelijke zone van de lymfeklier.
Rijst. 20. Lichtmicrofoto van de medulla van een lymfeklier. Hematoxyline-eosine kleuring. De lichte cerebrale sinussen worden begrensd door donkere cerebrale strengen die antigeen-gestimuleerde B-lymfocyten en hun effectorcellen - plasmacyten bevatten.
Tussen de lagen reticulo-endotheliocyten die enerzijds de lymfatische follikels en hersenkoorden bedekken en anderzijds het bindweefselstroma (capsule en trabeculae) bevinden zich spleetachtige ruimtes, die de sinussen van de lymfeklier worden genoemd. Het sinussysteem omvat de marginale (gelegen tussen de capsule en de follikels), de perifolliculaire corticale sinussen (tussen de follikels en trabeculae), de cerebrale (tussen de koorden en trabeculae) en het portaal (in het gebied van de concave deel - de poort van de lymfeklier) sinussen. In het sinussysteem circuleert lymfe vanuit de marginale sinus, waar de afferente lymfevaten stromen, door de tussenliggende sinussen naar de gate sinus, van waaruit de lymfe zal stromen door het systeem van externe lymfevaten. In dit geval wordt de lymfe geklaard door fagocytose van vreemde deeltjes door kustmacrofagen; lymfe is verrijkt met immunocompetente T- en B-lymfocyten, geheugencellen en immunoglobulinen (antilichamen).
Rijst. 21. Lichtmicrofoto van een perifeer fragment van een lymfeknoop, x 400. Gekleurd met hematoxyline-eosine. Pijlen geven stromale cellen aan - reticuloendotheliocyten.
Rijst. 22. Lichtmicroscopie van de lymfeklier: A - algemeen plan van de structuur, x 30; B - lymfoïde follikel met een licht reactief centrum, x 200; C - hersenkoord omgeven door reticulaire stroma, x 200.
De werkingsmechanismen van de lymfeklier zorgen voor een nauwe relatie van al zijn structurele componenten. Kustcellen en typische macrofagen van de follikels fagocyteren vreemde deeltjes die met de lymfe door het sinussysteem van de lymfeknoop gaan. Tegelijkertijd worden, met de deelname van lysosomale enzymen van macrofagen, de antigenen van gefagocyteerde deeltjes omgezet van een corpusculaire vorm in een moleculaire vorm die in staat is een immuunrespons te induceren: lymfocytproliferatie, B-lymfocyten die in plasmacyten veranderen (antilichaamproducenten), T-lymfocyten in effectoren (T-killers) en T-cellen geheugen. Antigeen-geactiveerde B-lymfocyten van de follikels gaan naar de hersenkoorden, waar ze veranderen in plasmacellen - producenten van antilichamen. Geheugencellen komen de bloedbaan binnen: daaruit worden effectorcellen gevormd na secundair contact met het antigeen.
Rijst. 23. Licht microfoto van een lymfeklier follikel, x 400. Hematoxyline-eosine kleuring. Men kan grote dendritische cellen zien die antigeenafhankelijke reproductie van B-lymfocyten stimuleren.
Dendritische cellen van de follikels van de corticale stof zijn een soort macrofagen die in staat zijn om complexen van antilichamen met antigenen op hun oppervlak te fixeren. Contact met dendritische cellen B-lymfocyten gestimuleerd om antistoffen aan te maken. Interdigiterende cellen van de paracorticale zone scheiden biologisch actieve stoffen af die de proliferatie en rijping van T-lymfocyten stimuleren, waardoor ze in effectorcellen (T-killers) veranderen.
Rijst. 24. Lichtmicrofoto van een fragment van een lymfeklierfollikel, x 1000. Hematoxyline-eosinekleuring. Men ziet grote immunocyten - B-lymfoblasten, die zich in het kiemcentrum vermenigvuldigen onder invloed van de stimulerende werking van dendritische macrofagen.
Het verschijnen van lymfeklieren werd opgemerkt aan het einde van de tweede maand van embryonale ontwikkeling in de vorm van zones van lokale ophopingen van mesenchymale cellen rond de lymfevaten. Van de buitenste laag van het mesenchym worden een capsule en trabeculae gevormd, van de binnenste - het reticulaire stroma van de knopen. De verwijdering van lymfoblasten en lymfocyten uit het beenmerg zorgt voor de vorming van hersenkoorden en lymfatische follikels aan het einde van de vierde maand van embryogenese. Even later wordt de thymusafhankelijke paracorticale zone bevolkt en worden de lymfeklieren verrijkt met macrofagen. Aan het einde van de vijfde maand krijgen de lymfeklieren morfologische kenmerken die kenmerkend zijn voor een volwassen organisme. Ze voltooien hun vorming tijdens de eerste drie jaar van het leven van een kind. Reactieve centra in de follikels verschijnen tijdens de immunisatie van het lichaam in het levensproces en de vorming van zijn beschermende functies. BIJ oude leeftijd het aantal reactieve centra in de follikels van de lymfeklieren neemt af, de fagocytische activiteit van macrofagen neemt af, sommige van de knooppunten atrofiëren en ze worden vervangen door vetweefsel.
HEMOLYMFATISCHE KNOOPPUNTEN ( nodi lymfatische haemalis) is een speciaal soort lymfeklieren, in de sinussen waarvan geen lymfe, maar bloed circuleert, en die de functie vervullen van zowel lymfoïde als myeloïde hematopoëse. Bij mensen bevinden hemolymfeklieren zich in het perirenale weefsel, rond de abdominale aorta, minder vaak in het achterste mediastinum. In structuur lijken ze op typische lymfeklieren, maar ze worden gekenmerkt door kleinere afmetingen, zwakkere ontwikkeling van de hersenkoorden en corticale follikels. Met de leeftijd werd involutie van de hemolymfeknopen opgemerkt: de cortex en medulla worden vervangen door vetweefsel of los vezelig bindweefsel.
MILT ( milt, pandrecht) is een ongepaard orgaan in de buikholte. De milt heeft een langwerpige vorm, gelokaliseerd in het linker hypochondrium. De massa is 100-150 g, de afmetingen zijn 10x7x5 cm In de milt vindt reproductie en antigeenafhankelijke differentiatie van lymfocyten plaats, evenals de eliminatie van erytrocyten en bloedplaatjes die hun levenscyclus hebben voltooid. De milt vervult ook de functie van bloed- en ijzerdepot, produceert biologisch actieve stoffen (splenine, een erytropoëse-remmingsfactor) en is in de embryonale periode een universeel hematopoëtisch orgaan. De milt is bedekt met een bindweefselcapsule, waaruit partities groeien in het orgaan - trabeculae. Het kapsel en de trabeculae bevatten, naast het bindweefsel dat rijk is aan collageen en elastische vezels, bundels gladde myocyten en vormen het musculoskeletale apparaat van de milt. In het parenchym van de milt worden rode en witte pulp onderscheiden.
Rijst. 25. Lichtmicrofoto van een rattenmilt. Hematoxyline-eosine kleuring. Kwantitatief overheersende rode pulp op het felroze preparaat (oxyfiel), witte pulp - intens basofiel, is een opeenhoping van lymfocyten.
Rijst. 26.
De witte pulp maakt ongeveer 20% van de massa van het orgaan uit en wordt gevormd door lymfocyten, plasmacellen, macrofagen, dendritische en interdigiterende cellen, waarvan het reticulaire weefsel het raamwerk is. Bolvormige ophopingen van dit soort cellen worden lymfatische follikels (knobbeltjes) van de milt genoemd. De diameter van de follikels is 0,3-0,5 mm, ze zijn omgeven door een capsule van reticuloendotheelcellen.
Rijst. 27. Semi-schematische weergave van lichtmicroscopie van een fragment van de milt. Hematoxyline-eosine kleuring.
De lymfatische follikel van de milt (Malpighiaans lichaam) heeft vier zones: peri-arterieel, mantel, marginaal en ook een licht (reactief of germinaal) centrum. De reactieve centra van de lymfatische follikels van de milt en lymfeklier zijn identiek in structuur en functie van de formatie. Ze bevatten B-lymfoblasten, typische macrofagen, dendritische en reticulaire cellen. Het verschijnen van reactieve centra in de follikels is een reactie op antigene stimulatie. De periarteriële zone is een opeenhoping van T-lymfocyten rond de slagader van de lymfatische follikel, of, zoals het ook wordt genoemd, de centrale slagader van de milt. De peri-arteriële zone is verrijkt met interdigiterende cellen - macrofagen, die in staat zijn om complexen van antilichamen met antigenen op hun oppervlak te fixeren en de proliferatie en rijping van T-lymfocyten te veroorzaken. De peri-arteriële zone van de miltfollikels is een analoog van de thymus-afhankelijke paracorticale zone van de lymfeklieren. De donkere mantelzone wordt gevormd door compact gelegen kleine B-lymfocyten en een klein aantal T-lymfocyten, plasmacellen en macrofagen. De marginale zone - de plaats waar de witte pulp overgaat in de rode - wordt gevormd door B- en T-lymfocyten, macrofagen en wordt begrensd door poreuze sinusoïdale hemocapillairen. Na de rijping van lymfocyten gaan ze van het lichte centrum en de peri-arteriële zone naar de mantel en de marginale zones met de volgende uitgang in de bloedbaan.
Rijst. 28. Lichtmicrofoto van een fragment van het lichaam van Malpighian, x 400. Gekleurd met hematoxyline-eosine. In het midden van de follikel is een licht kiemcentrum zichtbaar, aan de periferie waarvan zich een centrale slagader bevindt.
De marginale zone, als gevolg van contacten met hemocapillairen, hoopt zich op uit het bloed een groot aantal van antigenen en speelt daarom een belangrijke rol bij de immunologische activiteit van de milt. Een groot aantal pulpa-arteriolen die uit de centrale slagader komen, verlaten de witte pulp, maar keren dan terug en stromen in de sinussen van de marginale zone rond de knoop. Een groot aantal macrofagen en reticulo-endotheelcellen langs de periferie van de follikel dienen voor fagocytose en verwijdering van antigeen afval uit het bloed. Dendritische cellen die zich hier bevinden, absorberen en verzenden antigenen naar immunologisch competente cellen (T- en B-lymfocyten), die de systemische circulatie in de witte pulp verlaten, precies vanuit de sinusoïdale haarvaten van de marginale zone van de follikel. Geactiveerde lymfocyten migreren naar het kiemcentrum van de knobbel, veranderen in immunoblasten (de zogenaamde blasttransformatie van lymfocyten vindt plaats), prolifereren en veranderen in effectorcellen. Deze laatste komen de rode pulp binnen, waar de plasmacellen clusters vormen in de vorm van Billrott-strengen en antilichamen produceren die in het bloed worden afgegeven. Geactiveerde T-lymfocyten verlaten de rode pulpa en keren terug naar de algemene bloedsomloop.
Rijst. 29. Thymus-afhankelijke en thymus-onafhankelijke zones van de milt. De opeenhoping van T-lymfocyten (lichtcellen) rond de slagaders die uit de trabeculae komen, vormt een thymusafhankelijke zone. De lymfatische follikel en het lymfoïde weefsel van de witte pulp eromheen is een thymus-onafhankelijke zone. Hier zijn B-lymfocyten (donkere cellen), macrofagen en folliculaire procescellen aanwezig.
Lymfatische peri-arteriële omhulsels zijn een langwerpige opeenhoping van lymfocyten, die in de vorm van klauwen de slagaders van de witte pulp bedekken en aan de ene kant doorgaan in de lymfatische follikels van de milt. In het centrale deel van de vagina, dichter bij het lumen van het vat, zijn B-lymfocyten en plasmocyten geconcentreerd, aan de periferie - T-lymfocyten.
De rode pulp, die ongeveer 80% van de massa van de milt uitmaakt, is een opeenhoping van bloedcellen die zich ofwel in de omgeving van de reticulaire cellen of in het systeem van de vasculaire sinussen van de milt bevinden. Gebieden met rode pulp die zich tussen de sinussen bevinden, worden pulpakoorden van de milt genoemd. Ze voeren de processen uit van transformatie van B-lymfocyten in plasmacellen, evenals monocyten in macrofagen. Miltmacrofagen zijn in staat om oude of beschadigde rode bloedcellen en bloedplaatjes te herkennen en te vernietigen. In dit geval wordt het hemoglobine van de vernietigde erytrocyten gebruikt en wordt het een bron van ijzer voor de synthese van bilirubine en transferrine. De moleculen van de laatste worden door macrofagen van het rode beenmerg aan de bloedsomloop onttrokken en gebruikt bij het proces van erytrocytneogenese.
Rijst. dertig. Lichtmicrofoto van een fragment van de rode pulp van de milt, x 1000. Gekleurd met hematoxyline-eosine. Talrijke macrofagen worden gezien omringd door bloedcellen. Bleek gekleurde procesvormige cellen - stromale elementen van de milt - reticulaire cellen.
Rijst. 31. Elektronenmicrofoto van een fragment van de rode pulp van de milt. Men kan de poreuze endotheelwand van de veneuze sinussen en de lymfoïde banden van Billrott daartussen zien.
Rijst. 32. Scanning elektronenmicroscopie. Fragment van de veneuze sinus van de milt. Je kunt de opening zien tussen de endotheliocyten, waarin de gevormde elementen van het bloed worden geperst. De letters geven aan: N - neutrofiel; M - macrofaag, L - lymfocyt. Macrofagen regelen de kwaliteit van cellen die terugkeren naar de bloedbaan.
Het vasculaire systeem van de milt heeft een aantal kenmerken die ervoor zorgen dat de functies van dit orgaan worden uitgevoerd. Dientengevolge komt de miltslagader de poorten van de milt binnen, die vertakt in een systeem van vertakkingen in de trabeculae van de milt, de trabeculaire slagaders. De trabeculaire slagaders verdelen zich in de slagaders van de witte pulp van de milt, waarrond lymfocyten zijn gegroepeerd en peri-arteriële lymfatische omhulsels en follikels van de milt worden gevormd. Die delen van de witte pulpaslagaders die door de lymfatische follikels gaan, worden centrale slagaders genoemd, omdat ze dienen als centra voor de uitdrijving van lymfocyten tijdens de vorming van lymfatische follikels in ontogenese. De centrale slagaders gaan over in de slagaders van de rode pulpa, deze vallen uiteen in borstelarteriolen, die eindigen in ellipsoïde (mouw)arteriolen. Ellipsoïde arteriolen zijn omgeven door een soort "mouwen" - koppelingen van clusters van reticulaire cellen en reticulaire vezels die de rol spelen van arteriële sluitspieren van de milt. Via een systeem van hemocapillairen communiceren ellipsoïde arteriolen met de veneuze sinussen van de milt van een poreus type. Dit is het zogenaamde systeem van gesloten (gesloten) bloedcirculatie van de milt. Sommige van de haarvaten kunnen echter direct in de rode pulp openen en een open (open) miltcirculatiesysteem vormen. Veneuze sinussen met een aanzienlijke bloedvulling kunnen dienen als bloeddepot. Vanuit de veneuze sinussen stroomt bloed in de aderen van de rode pulp, vervolgens in de trabeculaire aderen en van de laatste in de miltader. In de wand van de veneuze sinussen in het gebied van hun overgang nabij de ader van de rode pulp, is er een opeenhoping van gladde myocyten die de veneuze sluitspieren van de milt vormen.
Rijst. 33. Schema van bloedtoevoer naar de milt Trabeculaire slagaders → pulpslagaders → arteriolen en haarvaten van de follikel → sinussen van de marginale zone → uitgang van T- en B-lymfocyten uit het vaatbed. Follikelarteriolen → borstelarteriolen van de rode pulp → sinusoïde haarvaten.
Rijst. 34. Sinusoïden in de rode pulp van de milt. Volgens de theorie van open circulatie (hierboven) komt bloed uit de haarvaten de rode pulp binnen en vervolgens in de sinusoïden. Volgens de theorie van de gesloten circulatie (van onderaf) openen de haarvaten direct in sinusoïden.
Rijst. 35. Lichtmicroscopie van miltfragmenten met verschillende histologische kleuringen: A - impregnatie met zilvernitraat (reticulair stroma is zichtbaar); B - kleuring met hematoxyline-eosine (structurele componenten van het orgelparenchym worden gevisualiseerd); C – kleuring met ijzerhematoxyline (verschillende dichtheid van lymfoïde elementen in de witte pulp is duidelijk zichtbaar).
Met de samentrekking van de veneuze sluitspieren hoopt het bloed zich op in de sinussen, het wordt dikker als gevolg van de impregnatie van het plasma door de wand van de veneuze sinussen. Bij gelijktijdige samentrekking van arteriële en veneuze sluitspieren wordt bloed in de milt afgezet. Ontspanning van arteriële en veneuze sluitspieren met gelijktijdige samentrekking van gladde myocyten van het kapsel en trabeculae van de milt bepaalt vooraf de afgifte van afgezet bloed in het veneuze bed.
Het leggen van de milt wordt uitgevoerd aan het begin van de tweede maand van de embryonale ontwikkeling in de vorm van ophopingen van mesenchymale cellen doordrongen van bloedvaten in het dorsale mesenterium. Uit het mesenchym wordt reticulair weefsel gevormd, dat laatste wordt bevolkt door bloedstamcellen. In de derde maand van embryogenese differentieert de peri-arteriële thymus-afhankelijke zone in de milt, in de vijfde maand worden reactieve centra en marginale zones van follikels gevormd, in de zesde maand kan rode pulp worden onderscheiden. Tegelijkertijd (van de derde tot de vijfde maand van de embryogenese) nemen de verschijnselen van myeloïde hematopoëse toe in de milt, het vervult de functies van een universeel hematopoëtisch orgaan. Vanaf de zesde maand tot de geboorte van een kind vervagen de manifestaties van myeloïde hematopoëse, ze worden vervangen door de processen van lymfocytopoëse. Op volwassen leeftijd vertoont de milt aanzienlijke herstellende vermogens; experimenteel bewezen de mogelijkheid van vernieuwing met het verlies van 80-90% van het parenchym. De massa van de milt neemt iets af op de leeftijd van 20 tot 30 jaar; in het interval van 30 tot 60 jaar is het stabiel. Op oudere leeftijd werden atrofie van de rode en witte pulp, proliferatie van bindweefselstroma, een afname van het gehalte aan macrofagen en lymfocyten onder parenchymale elementen, een toename van het gehalte aan granulocyten en weefselbasofielen en het verschijnen van megakaryocyten opgemerkt. Het gebruik van ijzer uit in de milt vernietigde erytrocyten gaat achteruit.
Intercellulaire interacties
bij het leveren van de immuunafweer van het lichaam.
Voor een adequate reactie op lichaamsvreemde stoffen (antigene stimulatie) is interactie en samenwerking van verschillende typen cellen van het immuunsysteem noodzakelijk. Onder hen worden cellen van macrofagen onderscheiden - monocyten bloed, histiocyten- bindweefselmacrofagen, beenmerg, peritoneale, alveolaire macrofagen, Langerhans-cellen huid, Kashchenko-Hofbauer-cellen placenta, stellate reticuloendotheliocyten lever, dendritisch en interdigiterende cellen lymfeklieren en milt, osteoclasten botweefsel microgliocyten zenuwstelsel. Er is een groep zogenaamde microfagen, waartoe ze behoren neutrofiele granulocyten bloed, evenals cellen die, onder bepaalde omstandigheden van functioneren, fagocytische eigenschappen kunnen vertonen, - endotheliocyten, fibroclasten. Ten slotte combineert de derde groep cellen verschillende populaties van T- en B-lymfocyten ( T-killers, T-helpers, T-suppressors, plasmacellen, T- en B-geheugencellen). De totale massa cellen die direct zorgen voor de immuunafweer van het lichaam is ongeveer 1% van het lichaamsgewicht.
Allereerst reageren T-helpers op het binnendringen van vreemde deeltjes in het lichaam: antigene determinanten binden aan specifieke receptoren op hun oppervlak. Het gevormde antigeenreceptorcomplex komt los van het oppervlak van het T-helperplasmolema en wordt gefixeerd door de oppervlaktereceptoren van de macrofaag. In het volgende stadium worden door macrofagen gemodificeerde antigenen overgebracht naar B-lymfocyten, die, onder invloed van antigene stimulatie en de activerende werking van T-helpers, in plasmacellen veranderen. Deze laatste synthetiseren eiwitmoleculen van immunoglobulinen (antilichamen), die selectief binden aan antigenen en hun inactivatie vooraf bepalen. T-helpers produceren, na contact met het antigeen, speciale chemicaliën die de proliferatie van T-killers stimuleren. Deze laatste hebben het vermogen om de celmembranen van bacteriën en cellen die antigene determinanten op hun oppervlak dragen, te vernietigen.
In elk van deze stadia kan gedeeltelijke inactivering van vreemd materiaal optreden, evenals de modificatie en overdracht naar andere celpopulaties om een immuunrespons te ontwikkelen. Het is mogelijk dat een antigeenbevattend deeltje wordt herkend en afgevoerd door een macrofaag zonder de deelname van een T-lymfocyt, wordt gesplitst door zijn lysosomale enzymen, en de resulterende antigene fragmenten worden overgebracht naar T- en B-lymfocyten en stimuleren hun transformatie in effectorcellen (T-killers en plasmacellen), evenals geheugencellen.
Interactie van cellen in de immuunrespons
De immuunrespons is mogelijk als gevolg van de activering van lymfocytklonen en bestaat uit twee fasen. In de eerste fase activeert het antigeen die lymfocyten die het herkennen. In de tweede (effector)fase coördineren deze lymfocyten de immuunrespons gericht op het elimineren van het antigeen.
humoristisch immuun antwoord
De humorale immuunrespons omvat macrofagen (antigeenpresenterende cellen), T-helpers en B-lymfocyten.
macrofaag absorbeert het antigeen dat het lichaam binnendringt en onderwerpt het aan verwerking - splitsing in fragmenten. Antigeenfragmenten worden samen met het MHC-molecuul op het celoppervlak blootgesteld. Het complexe "antigeenmolecuul MHC klasse II" wordt aangeboden aan T-helper (Fig. 35).
Rijst. 36. Antigeenherkenning door T-lymfocytreceptor. Met behulp van de T-lymfocytreceptor herkent de T-cel het antigeen, maar alleen in complex met het MHC-molecuul. In het geval van de T-helper is zijn molecuul, CD4, betrokken bij het proces, dat zich met zijn vrije uiteinde aan het MHC-molecuul bindt. Het antigeen dat door de T-cel wordt herkend, heeft twee plaatsen: de ene interageert met het MHC-molecuul, de andere (epitoop) bindt aan de T-lymfocytreceptor. Een soortgelijk type interactie, maar met de deelname van het CD8-molecuul, is kenmerkend voor het proces van herkenning door de T-killer van het antigeen dat is geassocieerd met het MHC klasse I-molecuul.
T-helper herkent het antigeen-MHC klasse II-molecuulcomplex op het oppervlak van de antigeenpresenterende cel. Het herkenningsproces omvat de interactie van het T-celreceptor-CD3-complex, dat zorgt voor specificiteit en deelname van ondersteunende co-stimulerende moleculen. Om de T-helper te activeren, is specifieke herkenning door de T-helper van een antigeenfragment op het oppervlak van een antigeenpresenterende cel onvoldoende. Activering van T-helpers wordt verschaft door de interactie van het B7-molecuul (CD80) op het oppervlak van de antigeenpresenterende cel met het CD28-molecuul op het oppervlak van de T-helper. T-helpers zijn bijzonder gevoelig voor stimulatie via CD28, die differentiëren tot T-helper 2 en B-cellen activeren via CD80. Bij zwakke expressie van CD28 en in aanwezigheid van het CTLA-molecuul ( C ytotoxischT - ik ymfocytp rotin) T-helpers 1 worden gevormd.
Herkenning door T-helper van de gewenste moleculen op het oppervlak van de antigeenpresenterende cel stimuleert de secretie van IL1. Geactiveerde IL1 T-helper synthetiseert IL2- en IL2-receptoren, waardoor de agonist de proliferatie van T-helpers en cytotoxische T-lymfocyten stimuleert. In het geval van T-helper hebben we het over autocriene stimulatie, wanneer de cel reageert op het middel dat zelf synthetiseert en afscheidt. Dus, na interactie met een antigeenpresenterende cel, verwerft T-helper het vermogen om te reageren op de werking van IL2 met een uitbarsting van proliferatie. De biologische betekenis van dit proces is de accumulatie van een dergelijke hoeveelheid T-helpers, die zal zorgen voor de vorming in de lymfoïde organen van het vereiste aantal plasmacellen die in staat zijn antilichamen tegen dit antigeen te produceren.
B-lymfocyt. B-lymfocytactivering omvat directe interactie van het antigeen met een immunoglobuline (Ig) op het oppervlak van de B-cel. In dit geval verwerkt de B-lymfocyt zelf het antigeen en presenteert zijn fragment in verband met het MHC II-molecuul op zijn oppervlak. Dit complex herkent een T-helper geselecteerd door hetzelfde antigeen dat betrokken was bij de selectie van deze B-lymfocyt. Twee paar moleculen zijn betrokken bij de activering van B-cellen: enerzijds de specifieke interactie van het antigeen met de Ig M-receptor op het oppervlak van de B-lymfocyt en anderzijds het CD40-molecuul op het oppervlak van de B-cel interageert met het CD40L (CD154) molecuul op het oppervlak van de T-helper, waardoor de B-cel wordt geactiveerd. Herkenning door de T-helperreceptor van het “antigen-molecuul MHC klasse II”-complex op het oppervlak van de B-lymfocyt leidt tot de secretie van IL2, IL4, IL5 en γ-IFN uit de T-helper, onder invloed van waarbij de B-cel wordt geactiveerd en prolifereert, waardoor een kloon wordt gevormd. De geactiveerde B-lymfocyt differentieert tot een plasmacel: het aantal ribosomen neemt toe, het granulaire endoplasmatisch reticulum en het Golgi-complex worden meer uitgesproken.
Plasma cel synthetiseert Ig. IL6 uitgescheiden door geactiveerde T-helpers stimuleert de secretie van Ig. Een deel van rijpe B-lymfocyten circuleert na antigeenafhankelijke differentiatie in het lichaam als geheugencellen.
mobiel immuun antwoord
De cellulaire immuunrespons wordt gekenmerkt door de proliferatie van toegewijde immunocompetente cellen die reageren met het antigeen in combinatie met het molecuul MHC klas l op het oppervlak van vreemde cellen of endogene antigenen in combinatie met het MHC klasse I-molecuul op het oppervlak van de eigen met virus geïnfecteerde en getransformeerde (tumor)cellen. De cytotoxische T-lymfocyt is betrokken bij de cellulaire immuunrespons.
reacties celgemedieerd cytolyse. Effectorcellen gebruiken hun receptoren om de doelcel te herkennen en te vernietigen. Niet alleen T-lymfocyten, maar ook andere subpopulaties van lymfoïde cellen en, in sommige gevallen, myeloïde cellen zijn verantwoordelijk voor celgemedieerde cytolyse. Het herkenningsproces omvat verschillende moleculen die worden blootgesteld aan het oppervlak van interagerende cellulaire partners:
√ specifieke antigenen, bijvoorbeeld virale peptiden op het oppervlak van geïnfecteerde cellen, in combinatie met het MHC-molecuul, worden herkend door cytotoxische T-celreceptoren, voornamelijk CD8+- en sommige subpopulaties van CD4+-cellen;
√ antigene determinanten van tumorcellen worden herkend door NK-cellen zonder deelname van MHC klasse I-molecuul;
√ antigeen-geassocieerde AT op het oppervlak van doelcellen wordt herkend door receptoren van Fc-fragmenten van NK-cellen (fenomeen van AT-afhankelijke cytotoxiciteit).
cytotoxisch T-lymfocyt(TC). Het antigeen dat op het oppervlak van de doelcel wordt gepresenteerd in een complex met het klasse I MHC-molecuul, bindt aan de cytotoxische T-lymfocytreceptor. Bij dit proces is het CD8-molecuul van het celmembraan Tc betrokken. IL2 uitgescheiden door T-helpers stimuleert de proliferatie van cytotoxische T-lymfocyten.
Verwoesting cellen-doelen. De cytotoxische T-lymfocyt herkent de doelcel en hecht eraan. Kleine korrels zijn aanwezig in het cytoplasma van een geactiveerde cytotoxische T-lymfocyt. Ze bevatten het cytolytische eiwit perforine. De perforinemoleculen die vrijkomen door de T-killer worden gepolymeriseerd in het membraan van de doelcel in aanwezigheid van Ca2+. De perforineporiën gevormd in het plasmamembraan van de doelcel laten water en zouten door, maar geen eiwitmoleculen. Als polymerisatie van perforine plaatsvindt in de extracellulaire ruimte of in het bloed, waar een overmaat aan calcium aanwezig is, dan zal het polymeer niet in staat zijn het membraan binnen te dringen en de cel te doden. De T-killer zelf is beschermd tegen het cytotoxische effect van perforine.
In praktisch werk worden de volgende histologische preparaten voor studie aangeboden:
1. SMEER VAN ROOD BEENMERG.
Kleuring: volgens Romanovsky-Giemsa (azuur II, eosine).
Vind bij lage en dan bij hoge vergroting van de microscoop sinusoïdale capillairen in het preparaat. In hun lumen zijn erytrocyten en leukocyten zichtbaar. Tussen de sinusoïdale haarvaten bevindt zich een reticulair weefsel, in de lussen waarvan hematopoëtische cellen zich in verschillende stadia van rijping bevinden. Cellen van de erytropoëtische reeks: a) proerytroblasten - grote cellen (15 micron) met een grote ronde kern, waarin de nucleoli duidelijk zichtbaar zijn. Een sterk basofiel cytoplasma omringt de kern met een smalle strook; b) basofiele erytroblasten - cellen (10-12 micron) met basofiel cytoplasma. De kern is rond, bevat meer heterochromatine en heeft een donkere kleur; c) polychromatofiele erytroblasten - zelfs kleinere cellen (8-10 micron), hun cytoplasma wordt gelijktijdig gekleurd met zowel zure als alkalische kleurstoffen en ziet er grijsachtig groenachtig uit. De kernen zijn intens gekleurd en hebben geen nucleoli; d) oxyfiele erytroblasten (normoblasten) - het cytoplasma van cellen bevat al een aanzienlijke hoeveelheid hemoglobine, daarom kleurt het oxyfiel, de kernen worden gepycnotiseerd, vaak gelyseerd en verdwijnen; e) reticulocyten - jonge erytrocyten. In het cytoplasma van deze cellen kan men granulaire mesh-structuren zien - de overblijfselen van de kern en het organel; g) rijpe erytrocyten - niet-nucleaire bloedcellen, met een homogeen oxyfiel cytoplasma.
Cellen van de granulocytopoëtische reeks: a) promyelocyten - grote cellen met grote ronde en lichte kernen. Het cytoplasma is matig basofiel, het bevat enkele azurofiele korrels (lysosomen); b) myelocyten bevatten specifieke granulariteit in het cytoplasma: eosinofiele hebben grote helderrode eosinofiele korrels; basofiel - donkerblauwe basofiele korrels; neutrofiel - hebben twee soorten kleine korrels die zowel basische als zure kleurstoffen waarnemen. Myelocyten rijpen tot metamyelocyten. Een kenmerkend morfologisch kenmerk van deze overgangsvorm van cellen is een afname van het volume van het cytoplasma en de kern, die de vorm aannemen van een gebogen staaf of hoefijzer. Afhankelijk van de korrels in het cytoplasma, worden metamyelocyten ook verdeeld in basofiel, oxyfiel en neutrofiel. In de loop van de rijping worden de kernen van metamyelocyten gesegmenteerd en veranderen de cellen in rijpe gesegmenteerde eosinofielen, basofielen en neutrofielen.
In een uitstrijkje van rood beenmerg zijn megakaryocyten gelokaliseerd in de buurt van de sinusoïden - zeer grote (> 50 m) cellen met verschillende polyploïde kernen en basofiel cytoplasma.
Onderzoek het monster, teken en label: 1. Proerytroblasten. 2. Polychromatofiele erytroblasten. 3. Neutrofiele metamyelocyten. 4. Megakaryocyten. 5. Rijpe bloedcellen (erytrocyten, leukocyten) in de sinusoïdale hemocapillair.
Voor wat morfologische kenmerken Kun je een polychromatofiele proerytrocyt van een erytrocyt onderscheiden?
In welk stadium van myelocytdifferentiatie kunnen basofiele, oxyfiele en neutrofiele cellen worden geïdentificeerd?
Welke cel is in een rood beenmerguitstrijkje groter: een megakaryoblast of een megakaryocyt?
Benoem op basis van de studie van een uitstrijkje van rood beenmerg de cellen die qua grootte overeenkomen met erytrocyten.
In welk stadium van differentiatie verliest de myelocyt zijn vermogen om te delen?
2. GOIMER (Thymus).
Bij lage vergroting is de lobvormige structuur van de klier zichtbaar. Elke lobule heeft een donkerder perifeer deel - de corticale substantie en een lichter binnenste deel - de medulla. Bij hoge vergroting, in het middelste deel van de medulla, zijn de epitheellichamen van Hassall zichtbaar - een concentrische gelaagdheid van epithelioreticulocyten. De stromale elementen van de lobulus zijn epithelioreticulocyten, die in contact komen met hun processen en een netwerk vormen. In de intervallen tussen de epithelioreticulocyten bevinden zich parenchymale elementen: T-lymfocyten (thymocyten) en macrofagen.
Onderzoek het monster, teken en label: 1. Klierlobule: 1. Cortex: a) T-lymfocyten; 2. Beenmerg: b) epithelioreticulocyten; c) Hassalls lichamen. II. Interlobulair bindweefsel. 3. Bloedvaten.
Maak op basis van het onderzoek van het thymuspreparaat een conclusie over de intensiteit van de mitosen in het subcapsulaire gebied, de cortex en de medulla.
Wat zijn de lichamen van Hassall en waar zijn ze gelokaliseerd in de thymus?
Is het epitheel dat het stroma van de thymus vormt eenlagig of meerlagig? Leg uit waarom.
3. LYMFE KNOOP.
Met een kleine vergroting van de microscoop is te zien dat de lymfeklier is bedekt met een bindweefselcapsule, van waaruit dunne partities, trabeculae, zich naar binnen uitstrekken. Tussen de trabeculae bevindt zich reticulair weefsel dat is geïnfiltreerd door talrijke lymfocyten. Lymfocyten zijn geconcentreerd langs de periferie van de knoop in de vorm van grote clusters van afgeronde vorm - follikels, die de corticale substantie van de lymfeknoop vormen. Van de follikels naar de diepte van de knoop vertrekken de hersenkoorden die de medulla vormen. Lichte ruimtes gevuld met reticulair weefsel en een klein aantal lymfocyten zijn sinussen. De marginale sinus, gelegen tussen de follikels en de capsule, gaat over in de intermediaire corticale sinussen, die op hun beurt doorgaan in de tussenliggende cerebrale sinussen, die lymfe verzamelen in de centrale sinus bij de poort van de lymfeklier.
Maak een schets van het preparaat en benoem: 1. Capsule. 2. Trabeculae. 3. Cortex. 4. De medulla. 5. Follikels. 6. Hersenbanden. 7. Marginale sinus. 8. Tussenliggende corticale sinussen. 9. Tussenliggende cerebrale sinussen. 10. Poort van de lymfeklier. 11. Reticulair weefsel.
Trek op basis van het bestudeerde preparaat een conclusie over de antigene stimulatie van de lymfeklier en licht je antwoord toe.
Wat circuleert er in de sinussen van de lymfeklier?
Geef op het preparaat de plaats aan waar zich de meeste plasmacellen bevinden.
4. MILT.
Gekleurd met hematoxyline en eosine.
Bij een lage vergroting van de microscoop is duidelijk een dicht bindweefselkapsel te zien, van waaruit de trabeculae zich uitstrekken tot in het orgaan. Het kapsel van de milt is bedekt met mesothelium en bevat een kleine hoeveelheid gladde spiercellen, die, door samen te trekken, bijdragen aan de afgifte van bloed door de poorten van het orgel. Tussen de trabeculae bevindt zich de witte en rode pulp van de milt. De witte pulp bestaat uit lymfatische follikels die een centrale slagader hebben. De rode pulp wordt gevormd door reticulair weefsel, bloedcellen, voornamelijk erytrocyten en veneuze sinussen.
Onderzoek het preparaat met een sterke vergroting, teken een klein gebied en wijs aan: 1. Miltcapsule. 2. Trabeculae. 3. Witte pulp (lymfatische follikels): a) centrale slagader; b) kiemcentrum. 4. Rode pulpa: a) reticulair weefsel, b) erytrocyten; c) leukocyten. 5. Veneuze sinus.
Wat zijn de belangrijkste onderscheidende kenmerken van de structuur van de lymfatische follikels van de milt en lymfeklieren.
Maak een conclusie waar er meer lymfoïde knobbeltjes zijn met voortplantingscentra in de milt of lymfeklier en leg uit waarom?
DEMOPRODUCTEN:
1. Een uitstrijkje van het rode beenmerg van een 5-jarig kind.
2. Thymusklier van een 12-jarig kind.
3. Reticulaire vezels in de lymfeklier. Zilver impregnatie.
ELEKTRONISCHE MICROFOTOGRAFIE:
1. Rode beenmergcellen.
2. Epithelioreticulocyten van de thymus.
3. T-lymfocyten.
4. Sinus van de lymfeklier.
5. Sinus van de milt.
Situationele taken:
1. Bij de voorbereiding is er een snee in het buisvormige bot van een kind van 3-5 jaar oud, een jonge man van 12-18 jaar oud en een oud persoon. Hoe verandert de toestand en topografie van het rode beenmerg met de leeftijd?
2. De thymus is verwijderd bij een pasgeboren kind. Als gevolg van deze operatie nam haar vermogen om antistoffen aan te maken sterk af. Leg de reden voor dit fenomeen uit.
3. Er zijn microfoto's van de lymfeklieren van het buikvlies, gefotografeerd op het hoogtepunt van de spijsvertering en in rust. Hoe kan men tijdens de spijsvertering een lymfeklier onderscheiden en dit fenomeen verklaren?
4. Het dier werd direct na de geboorte onder steriele omstandigheden geplaatst. Kunnen zich in deze situatie secundaire follikels vormen? lymfeklieren zo ja, waarom niet, zo nee, waarom niet?
5. In de oudheid werd bij marathonlopers de milt verwijderd. Leg uit waarom?
Lezingenplan:
Classificatie, functies en algemeen principe structuren van de hematopoëtische organen
Het concept van lymfoïde en myeloïde weefsel, de ontwikkeling van myeloïde hematopoëse
Rood beenmerg (RMB):
3.1. KKM-functies
3.2. De structuur van KKM
3.3. Kenmerken van de bloedtoevoer van de KCM
3.4. Regeneratie van KKM
Thymus:
4.1. Thymus functies
4.2. thymus ontwikkeling
4.3. De structuur van de thymus
4.4. Kenmerken van de bloedtoevoer naar de thymus. Hemato-thymus barrière.
4.5. Leeftijdskenmerken van de thymus
Functies van de hematopoëtische organen
De organen van hematopoëse en immuunafweer vormen een enkel systeem met bloed en lymfe, dat:
Zorgt voor een continu proces van vernieuwing van bloedcellen als gevolg van constante proliferatie en differentiatie van cellen in overeenstemming met de behoeften van het lichaam.
Creëert en implementeert een complex van beschermende reacties tegen de schadelijke effecten van externe en interne omgevingsfactoren, immuunbewaking van de activiteit van de cellen van uw lichaam.
Ondersteunt de integriteit en individualiteit van het organisme dankzij het vermogen van de cellen van het immuunsysteem om de structurele componenten van het lichaam van de alien te onderscheiden en deze te vernietigen.
De organen van hematopoëse en immunogenese omvatten:
Rood beenmerg (RMB),
Lymfe- en hemolymfeklieren,
Milt,
Lymfoïde formaties spijsverteringsstelsel, waaronder amandelen, Peyer's patches, appendix, lymfoïde formaties van de reproductieve, respiratoire, excretiesystemen.
Alle organen van hematopoëse en immunogenese zijn onderverdeeld in: centraal en perifere.
Tot staat centraal KCM en thymus. Hematopoëtische stamcellen zijn daarin gelokaliseerd en de eerste fase van lymfocytdifferentiatie vindt plaats, genaamd antigeen-onafhankelijk.
Tot perifere organen omvatten: milt, lymfatische en hemolymfeknopen, lymfoïde formaties langs de spijsverteringsbuis, genitale, respiratoire, uitscheidingssystemen. Deze organen voeren antigeen-afhankelijk differentiatie van lymfocyten.
Het algemene principe van de structuur van de hematopoëtische organen
De basis van alle vormen van hematopoëtische organen stroma de component vertegenwoordigd door reticulair weefsel, de enige uitzondering is de thymus, de stromale component wordt vertegenwoordigd door epithelioreticulair weefsel van epitheliale oorsprong. Stromacellen voeren ondersteunende, trofische en regulerende functies uit die in elk orgaan aanwezig zijn karakteristieke kenmerken. Ze creëren een speciale micro-omgeving, synthetiserend hematopoëtinen voor de juiste ontwikkeling van hematopoëtische cellen zijn GAG's zuur en neutraal, evenals het eiwit laminine, dat een driedimensionaal netwerk creëert voor de migratie van bloedcellen.
Alle organen van hematopoëse en immunogenese onder stromale cellen bevatten een groot aantal macrofagen, die betrokken zijn bij de rijping en differentiatie van gevormde gevormde elementen, evenals bij de fagocytose van vernietigde cellen, die deelnemen aan hun gebruik.
Het stroma van de hematopoëtische organen bevat vasculaire component, die wordt weergegeven door speciale bloedvaten, sinuscapillairen, met hoog endotheel, die op zijn beurt zorgt voor de herkenning van rijpe cellen, ze kan sorteren en de migratie van gevormde elementen naar de bloedbaan verzekert.
In het netwerk van stroma-creërend weefsel bevinden zich bloedcellen in verschillende stadia van rijping - hematopoëtische component.
Het concept van lymfoïde en myeloïde weefsel, de ontwikkeling van myeloïde hematopoëtische organen
Hematopoëtische cellen Samen met het stroma vormen ze twee soorten weefsels, myeloïde en lymfoïde:
Myeloïde weefsel- dit is een reticulair weefsel, met zich daar ontwikkelende cellen van de myeloïde reeks ( erytropoëse, trombopoëse, granulocytopoëse, monocytopoëse ) en lymfoïde (B-lymfocytopoëse). Myeloïde weefsel vormt de basis van myeloïde hematopoëtische organen, waaronder bij mensen rood beenmerg.
Lymfoïde weefsel- dit is een reticulair of epithelioreticulair weefsel (thymus), waarin zich cellen van de lymfoïde reeks bevinden ( lymfocytopoëse) in verschillende stadia van ontwikkeling. Lymfoïde weefsel vormt de organen van lymfoïde hematopoëse, waaronder: thymus, milt, lymfe- en hemolymfeknopen en lymfoïde elementen in de wand van verschillende organen en systemen.
Ontwikkeling van myeloïde hematopoëse:
Er zijn drie perioden in ontwikkeling:
mesoblastisch
Hepatolienal
medullair
Mesoblastisch (2 weken - 4 maanden): de eerste bloedcellen worden gevonden in het 13-19 dagen oude embryo in het mesoderm van de dooierzak. Intravasculair differentieert een deel van de bloedstamcellen tot erytroblasten (grote cellen met een kern). Extravasculaire granulocyten worden gevormd: neutrofielen en eosinofielen. De activiteit van mesoblastische hematopoëse neemt af in de 6e week en eindigt bij de 4e maand van embryogenese.
Hepatolienal (2 maanden - 7 maanden): in de lever begint hematopoëse na 5-6 weken en bereikt een maximum tegen de 5e maand van embryogenese. Alle gevormde elementen zijn erytrocyten en bloedplaatjes worden tijdens deze periode extravasculair gevormd. Tegen de tijd van geboorte kunnen enkele foci van hematopoëse in de lever achterblijven. In de milt worden foci van myeloïde hematopoëse gevonden vanaf de 20e week van embryogenese, foci van lymfoïde hematopoëse verschijnen iets later en vanaf de 8e maand van embryogenese blijft alleen lymfoïde hematopoëse erin.
Medullair of medullair: begint parallel met de ontwikkeling van het botskelet en gaat door gedurende het hele leven. Cellen van twee typen beginnen te groeien en differentiëren in de holte van het primaire bot: vanaf 2 maanden, mechanoblasten (vormen reticulair weefsel dat alle botholten vult) en vanaf 3 maanden - bloedstamcellen, die eilanden van hematopoëse vormen. Tegen de 4e maand van de embryogenese wordt de BMC het belangrijkste hematopoëtische orgaan en vult het de holtes van platte en buisvormige botten. Bij een 7-jarig kind wordt BCM in de diafyse van buisvormige botten bleek, geel beenmerg verschijnt en begint te groeien. Bij een volwassene wordt BMC alleen bewaard in de epifysen van buisvormige botten en in platte botten. Op oudere leeftijd krijgt het beenmerg (zowel rood als geel) een slijmerige consistentie en wordt het gelatineus beenmerg genoemd.
Morfologie van rood beenmerg (RMB)
rood beenmerg (mergossiumrubra) is het centrale orgaan van hematopoëse en immunogenese, dat een populatie van bloedstamcellen bevat en deelneemt aan de vorming van cellen van de myelocytische en lymfocytische reeks.
KKM-functies:
Hematopoëtisch - in het rode beenmerg ontstaan alle hematopoëtische kiemen op basis van een zichzelf in stand houdende populatie van bloedstamcellen
immuun- antigeen-onafhankelijke differentiatie van lymfocyten vindt plaats in het rode beenmerg
Regelgeving- hematopoëtines die vrijkomen in het rode beenmerg beïnvloeden de processen van hematopoëse in alle organen van hematopoëse, en gesynthetiseerde cytokinen reguleren de immunogenese.
Bij een volwassene is de massa van de CMC 1,5 - 2 kg, wat 4-5% van het lichaamsgewicht is. Het heeft een rode kleur en een semi-vloeibare consistentie. zijn basis of stromale component vormt reticulair weefsel, bestaande uit reticulaire procescellen (reticulocyten) en intercellulaire substantie die reticulaire vezels bevat. Het vormt niet alleen een driedimensionaal netwerk dat een ondersteunende functie vervult, maar zijn cellen synthetiseren hematopoëtische factoren, zonder welke hematopoëse niet wordt uitgevoerd. Reticulocyten die zich rond de wand van de bloedbijholten bevinden, worden genoemd adventieve cellen. Deze cellen kunnen samentrekken, waardoor de migratie van bloedcellen door de bloedvaten wordt vergemakkelijkt. Naast reticulocyten wordt de stromale component weergegeven door: adipocyten, macrofagen, net zoals endostale cellen(bindweefselbekleding van botholten) - osteoblasten en osteocyten.
Adipocyten bevinden zich op eilanden en leveren energie voor hematopoëse; vul het volume, creëer de druk die nodig is voor het functioneren van de sinussen, en produceer ook biologisch actieve stoffen die het volume van hematopoëtisch weefsel reguleren.
Macrofagen vervullen verschillende functies: scheiden groeifactoren af en fagocyteren cellen die de selectie niet hebben doorstaan; macrofagen die migreren vanuit de milt brengen componenten voor de synthese van hemoglobine, en botmacrofagen osteoclasten reguleren de grootte van botlacunes.
Osteoblasten en osteocyten, die een stijf skelet vormen, leveren sporenelementen die nodig zijn voor hematopoëse.
Tussen de reticulaire cellen bevinden zich een groot aantal holtes waarin hemocapillairen liggen. Ze zijn erg dun en geven kleur aan KKM. Rond de bloedcapillairen bevinden zich veel vrijliggende bloedcellen van myelocytische en lymfocytische rijen in verschillende stadia van differentiatie, evenals een zichzelf in stand houdende populatie van pluripotente stamcellen. De proliferatie in de RMC is zeer actief en creëert ongeveer 200 miljard cellen per dag.
De plaatsen waar de proliferatie en differentiatie van bloedcellen plaatsvindt, worden genoemd eilandjes van hematopoëse. Deze eilanden vormen over het algemeen hematopoëtische component.
Er zijn drie soorten eilanden:
erytropoëtisch het eilandje bevat een centraal gelegen macrofaag genaamd cel-oppas waarrond erytroïde cellen zich in verschillende stadia van ontwikkeling bevinden (van kolonievormende erytroïde cellen en erythroblast tot reticulocyt). Macrofaag maakt groeifactoren vrij sialoadhesins het houdt erytroïde cellen om zich heen en voorziet ze van ijzer vanwege de aanwezigheid in zijn cytoplasma transferrine, die 4 ijzeratomen bindt, produceren macrofagen ook erytropoëtine, vitamine D3 en fagocyteren de kernen die tijdens het rijpingsproces uit de erytrocyt worden uitgeworpen.
granulocytopoëtische eilandjes kunnen van drie soorten zijn, afhankelijk van welke granulocyten worden gevormd: neutrofiel, eosinofiel of basofiel, meestal zijn ze gelokaliseerd in de buurt van het endosteum. Elk eilandje is omgeven door een laag proteoglycanen, die een micro-omgeving creëert voor granulocytdifferentiatie. Naarmate ze rijpen, lost dit membraan op en de granulocyten, die amoeboïde bewegingen maken, migreren naar de sinussen en gaan de bloedbaan in.
trombocytopoëtische het eilandje bevindt zich in de buurt van de sinuscapillairen en omvat: megakaryocyten. Megakaryocyten zijn zeer grote cellen met reusachtige gelobde kernen. Het duwt de pseudopod tussen de endotheliocyten in de capillaire holte en de bloedstroom, deze gebieden komen los en veranderen in bloedplaatjes. Deze methode van losmaken van het cytoplasma wordt " clasmosis". Uit één megakaryocyt worden ongeveer 2000 bloedplaatjes gevormd.
Daarnaast zijn er drie categorieën lymfoïde cellen die rond de bloedvaten in de BMC liggen:
Stam lymfoïde cellen zonder receptoren.
voorgangers T - lymfocyten receptoren hebben en migreren naar de thymus.
voorgangers B - lymfocyten, waarbij een uniek proces van vorming van immunoglobuline-genen wordt uitgevoerd.
Bovendien ontwikkelen zich monocyten, toekomstige macrofagen, in RMC.
Om de processen van hematopoëse en immunogenese te laten plaatsvinden, zijn regulatoren nodig, die zijn onderverdeeld in -
Yoffey en Curtis (Yoffey, Courtice, 1970) combineerden de lymfoïde en hematopoëtische systemen tot een enkel lymfomyeloïde complex (Fig. B.6).
Het complex is een systeem van organen en weefsels, waarvan het parenchym cellen van mesenchymale oorsprong bevat. Het omvat: beenmerg, thymus, milt, lymfeklieren, intestinaal lymfoïde weefsel en bindweefsel.
Functionele cellen van het lymfoïde systeem worden vertegenwoordigd door lymfocyten, macrofagen, antigeenpresenterende cellen en, in sommige weefsels, epitheelcellen. Al deze cellen functioneren als onderdeel van geïsoleerde organen of diffuse formaties.
Lymfoïde organen worden geclassificeerd als primaire (centrale) of secundaire organen. De primaire lymfoïde organen zijn het rode beenmerg en de thymus.
Het functionele doel van het complex is om hematopoëse (myelopoëse) en de vorming van cellen van het immuunsysteem (lymfopoëse) te verzekeren. Onder de organen en weefsels van het complex bevinden zich echte lymfopoëse, waarin alleen lymfopoëse optreedt (thymus, lymfeklieren, intestinaal lymfoïde weefsel) en "gemengde" formaties, waar zowel lymfopoëse als myelopoëse aanwezig zijn (beenmerg, milt).
Het is in de primaire organen dat het repertoire van specificiteiten van lymfocyt-antigeen-herkennende receptoren wordt gevormd, en lymfocyten verwerven zo het vermogen om alle antigenen te herkennen die het lichaam tijdens het leven kan tegenkomen. Verder worden deze cellen geselecteerd op tolerantie (reactiviteit) voor auto-antigenen, waarna alleen vreemde antigenen worden herkend in perifere lymfoïde organen of formaties.
In de thymus 'leren' T-cellen bovendien hun eigen MHC-moleculen te herkennen. Het is echter bekend dat sommige lymfocyten zich buiten de primaire organen ontwikkelen.
Van de primaire organen migreren lymfocyten om hun functies uit te voeren langs de bloedbaan naar het perifere lymfoïde weefsel - lymfeklieren, milt en lymfoïde weefsel van de slijmvliezen (Peyer's patches, amandelen). Deze verplaatsing van lymfocyten van de centrale organen van het immuunsysteem naar de periferie is de belangrijkste migratieroute. Daarnaast is er een recyclingpad. Lymfatische vaten die het lichaam afvoeren, verzamelen extracellulaire vloeistof - lymfe - samen met lymfocyten verspreid over het lichaam en brengen deze naar de lymfeklieren. Na enige tijd in de lymfeklieren verzamelen de lymfocyten zich in de efferente efferente lymfevaten. Hiervan komen lymfocyten het belangrijkste lymfevat binnen - het thoracale kanaal, van waaruit ze weer terugkeren naar de bloedbaan via de linker subclavia-ader (Fig. 6.1 en Fig. 6.2).
Lymfocyten behoren dus tot de categorie cellen die wijd verspreid in het lichaam zijn. En in het lichaam van de mens en gewervelde dieren zijn ze gegroepeerd in drie soorten associaties (Fig. 6.14). Verschillende types organisatie van lymfocyten zorgt voor de meest effectieve manifestatie van het lymfoïde systeem wanneer het een vreemd antigeen tegenkomt.
De immuunrespons op antigenen die het lichaam binnenkomen via de slijmvliezen begint met de priming van lymfocyten, voornamelijk in de pleisters van Peyer.
Verschillende lymfoïde organen beschermen verschillende lichaamssystemen: de milt reageert op antigenen die in het bloed circuleren; lymfeklieren reageren op antigenen die door de lymfevaten komen; het lymfoïde weefsel van de slijmvliezen beschermt de slijmvliezen.
Lymfocyten zijn meestal geen bezonken, maar circulerende cellen; ze migreren constant van de bloedbaan naar de lymfoïde organen en komen weer in de bloedbaan terecht.
In vivo vinden de complexe cellulaire interacties die de basis vormen van de immuunrespons plaats in perifere of secundaire lymfoïde organen, waaronder de lymfeklieren, milt en ophopingen van diffuus lymfoïde weefsel in de slijmvliezen van de luchtwegen, spijsvertering en urogenitale traktaten.
Secundaire lymfoïde weefsels worden bevolkt door cellen van reticulaire oorsprong, evenals macrofagen en lymfocyten, waarvan de voorlopers beenmergstamcellen zijn. Stamcellen differentiëren tot immunocompetente T- en B-lymfocyten. In dit geval differentiëren T-lymfocyten tot immunocompetente cellen in de thymus en B-lymfocyten in het beenmerg. Vervolgens bevolken lymfocyten lymfoïde weefsels, waar de immuunrespons optreedt (Fig. 11: "Stamcellen (SC) van het beenmerg differentiëren in primaire lymfoïde organen tot immunocompetente T- en B-lymfocyten, die vervolgens secundaire lymfoïde organen bevolken"). (Lymfoïde weefsel geassocieerd met slijmvliezen, waarvan het beschermende effect is gebaseerd op de productie van IgA, wordt vaak aangegeven met de afkorting
Rijst. 17. Lokalisatie van plaatsen van lymfocytopoëse in het menselijk lichaam.
DE LYMFE KNOPEN ( nodi lympnatici) - een boonvormige verdikking langs de lymfevaten, waar antigeenafhankelijke reproductie van B- en T-lymfocyten plaatsvindt, hun verwerving van immuuncompetentie, evenals de zuivering van lymfe van vreemde deeltjes. De totale massa van lymfeklieren is 1% van het lichaamsgewicht, dat wil zeggen ongeveer 700 g. Lymfeklieren vormen meer dan 50 groepen. Volgens topografie zijn ze verdeeld in knooppunten van het lichaam (somatisch), ingewanden (visceraal) en gemengd, waarbij ze lymfe verzamelen van zowel de ingewanden als andere organen. De grootte van de lymfeklieren ligt in het bereik van 5-10 mm.
De lymfeklier is bedekt met een bindweefselcapsule, van waaruit bindweefselpartities zich uitstrekken tot in de orgel - trabeculae. In de capsule van sommige lymfeklieren werden gladde myocyten gevonden, die deelnemen aan de vorming van het bewegingsapparaat van de knoop.
Rijst. achttien. Diagram van de structuur van de lymfeklier. De rechterkant van de figuur toont de vascularisatie van de structurele elementen van het orgel.
Het parenchym van de knoop wordt gevormd door B- en T-lymfocyten, waarvan het skelet het reticulaire weefsel vormt. Er zijn corticale en medulla van de lymfeklier. De corticale substantie wordt gevormd door lymfatische follikels (knobbeltjes) die zich onder de capsule bevinden - bolvormige clusters van B-lymfocyten met een diameter van 0,5-1 mm. Naast B-lymfocyten omvat de samenstelling van de lymfeklierfollikels zowel typische macrofagen als een speciale variëteit van zichzelf, dendritische cellen genoemd. Uitwendig is de follikel bedekt met reticuloendotheliocyten - cellen die de morfologie van reticulaire cellen combineren met de functie van het endotheel, aangezien ze de sinussen van de lymfeklieren bekleden. Onder de reticuloendotheliocyten bevinden zich een aanzienlijk aantal vaste macrofagen, de zogenaamde kustcellen. Elke follikel bevat een licht (reactief of germinaal) centrum, waar lymfocyten zich vermenigvuldigen en waar B-lymfoblasten voornamelijk gelokaliseerd zijn, en een donkere perifere zone, waarin kleine en middelgrote lymfocyten compact zijn gelokaliseerd. Een toename van het aantal en de grootte van de reactieve centra van de follikels van de lymfeklieren duidt op antigene stimulatie van het lichaam.
Rijst. negentien. Lichtmicrofoto van een fragment van een lymfeknoop, x 200. Gekleurd met hematoxyline-eosine.
De medulla van de lymfeklier wordt gevormd door medullaire koorden - lintvormige clusters van B-lymfocyten, plasmacellen en macrofagen, langwerpig in de richting van de poort van de knoop naar de follikels. Uiterlijk zijn de hersenkoorden, evenals de follikels van de corticale substantie, bedekt met reticuloendotheliocyten. Tussen de hersenkoorden en follikels, respectievelijk, tussen de medulla en de cortex van de lymfeklier, is er een diffuse ophoping van T-lymfocyten, die de paracorticale zone wordt genoemd. Macrofagen in de paracorticale zone worden vertegenwoordigd door een verscheidenheid aan zogenaamde interdigiterende cellen die met elkaar in contact komen via vingervormige processen en stoffen produceren die de proliferatie van T-lymfocyten stimuleren. De corticale en medulla zijn dus bursa-afhankelijk en de paracorticale laag is de thymus-afhankelijke zone van de lymfeklier.
Rijst. 20. Lichtmicrofoto van de medulla van een lymfeklier. Hematoxyline-eosine kleuring. De lichte cerebrale sinussen worden begrensd door donkere cerebrale strengen die antigeen-gestimuleerde B-lymfocyten en hun effectorcellen - plasmacyten bevatten.
Tussen de lagen reticulo-endotheliocyten die enerzijds de lymfatische follikels en hersenkoorden bedekken en anderzijds het bindweefselstroma (capsule en trabeculae) bevinden zich spleetachtige ruimtes, die de sinussen van de lymfeklier worden genoemd. Het sinussysteem omvat de marginale (gelegen tussen de capsule en de follikels), de perifolliculaire corticale sinussen (tussen de follikels en trabeculae), de cerebrale (tussen de koorden en trabeculae) en het portaal (in het gebied van de concave deel - de poort van de lymfeklier) sinussen. In het sinussysteem circuleert lymfe vanuit de marginale sinus, waar de afferente lymfevaten stromen, door de tussenliggende sinussen naar de gate sinus, van waaruit de lymfe zal stromen door het systeem van externe lymfevaten. In dit geval wordt de lymfe geklaard door fagocytose van vreemde deeltjes door kustmacrofagen; lymfe is verrijkt met immunocompetente T- en B-lymfocyten, geheugencellen en immunoglobulinen (antilichamen).
Rijst. 21. Lichtmicrofoto van een perifeer fragment van een lymfeknoop, x 400. Gekleurd met hematoxyline-eosine. Pijlen geven stromale cellen aan - reticuloendotheliocyten.
Rijst. 22. Lichtmicroscopie van de lymfeklier: A - algemeen plan van de structuur, x 30; B - lymfoïde follikel met een licht reactief centrum, x 200; C - hersenkoord omgeven door reticulaire stroma, x 200.
De werkingsmechanismen van de lymfeklier zorgen voor een nauwe relatie van al zijn structurele componenten. Kustcellen en typische macrofagen van de follikels fagocyteren vreemde deeltjes die met de lymfe door het sinussysteem van de lymfeknoop gaan. Tegelijkertijd worden, met de deelname van lysosomale enzymen van macrofagen, de antigenen van gefagocyteerde deeltjes omgezet van een corpusculaire vorm in een moleculaire vorm die in staat is een immuunrespons te induceren: lymfocytproliferatie, B-lymfocyten die in plasmacyten veranderen (antilichaamproducenten), T-lymfocyten in effectoren (T-killers) en T-cellen geheugen. Antigeen-geactiveerde B-lymfocyten van de follikels gaan naar de hersenkoorden, waar ze veranderen in plasmacellen - producenten van antilichamen. Geheugencellen komen de bloedbaan binnen: daaruit worden effectorcellen gevormd na secundair contact met het antigeen.
Rijst. 23. Licht microfoto van een lymfeklier follikel, x 400. Hematoxyline-eosine kleuring. Men kan grote dendritische cellen zien die antigeenafhankelijke reproductie van B-lymfocyten stimuleren.
Dendritische cellen van de follikels van de corticale stof zijn een soort macrofagen die in staat zijn om complexen van antilichamen met antigenen op hun oppervlak te fixeren. Contact met dendritische cellen B-lymfocyten gestimuleerd om antistoffen aan te maken. Interdigiterende cellen van de paracorticale zone scheiden biologisch actieve stoffen af die de proliferatie en rijping van T-lymfocyten stimuleren, waardoor ze in effectorcellen (T-killers) veranderen.
Rijst. 24. Lichtmicrofoto van een fragment van een lymfeklierfollikel, x 1000. Hematoxyline-eosinekleuring. Men ziet grote immunocyten - B-lymfoblasten, die zich in het kiemcentrum vermenigvuldigen onder invloed van de stimulerende werking van dendritische macrofagen.
Het verschijnen van lymfeklieren werd opgemerkt aan het einde van de tweede maand van embryonale ontwikkeling in de vorm van zones van lokale ophopingen van mesenchymale cellen rond de lymfevaten. Van de buitenste laag van het mesenchym worden een capsule en trabeculae gevormd, van de binnenste - het reticulaire stroma van de knopen. De verwijdering van lymfoblasten en lymfocyten uit het beenmerg zorgt voor de vorming van hersenkoorden en lymfatische follikels aan het einde van de vierde maand van embryogenese. Even later wordt de thymusafhankelijke paracorticale zone bevolkt en worden de lymfeklieren verrijkt met macrofagen. Aan het einde van de vijfde maand krijgen de lymfeklieren morfologische kenmerken die kenmerkend zijn voor een volwassen organisme. Ze voltooien hun vorming tijdens de eerste drie jaar van het leven van een kind. Reactieve centra in de follikels verschijnen tijdens de immunisatie van het lichaam in het levensproces en de vorming van zijn beschermende functies. Op oudere leeftijd neemt het aantal reactieve centra in de follikels van de lymfeklieren af, de fagocytische activiteit van macrofagen neemt af, sommige van de knooppunten atrofiëren en ze worden vervangen door vetweefsel.
HEMOLYMFATISCHE KNOOPPUNTEN ( nodi lymfatische haemalis) is een speciaal soort lymfeklieren, in de sinussen waarvan geen lymfe, maar bloed circuleert, en die de functie vervullen van zowel lymfoïde als myeloïde hematopoëse. Bij mensen bevinden hemolymfeklieren zich in het perirenale weefsel, rond de abdominale aorta, minder vaak in het achterste mediastinum. In structuur lijken ze op typische lymfeklieren, maar ze worden gekenmerkt door kleinere afmetingen, zwakkere ontwikkeling van de hersenkoorden en corticale follikels. Met de leeftijd werd involutie van de hemolymfeknopen opgemerkt: de cortex en medulla worden vervangen door vetweefsel of los vezelig bindweefsel.
MILT ( milt, pandrecht) is een ongepaard orgaan in de buikholte. De milt heeft een langwerpige vorm, gelokaliseerd in het linker hypochondrium. De massa is 100-150 g, de afmetingen zijn 10x7x5 cm In de milt vindt reproductie en antigeenafhankelijke differentiatie van lymfocyten plaats, evenals de eliminatie van erytrocyten en bloedplaatjes die hun levenscyclus hebben voltooid. De milt vervult ook de functie van bloed- en ijzerdepot, produceert biologisch actieve stoffen (splenine, een erytropoëse-remmingsfactor) en is in de embryonale periode een universeel hematopoëtisch orgaan. De milt is bedekt met een bindweefselcapsule, waaruit partities groeien in het orgaan - trabeculae. Het kapsel en de trabeculae bevatten, naast het bindweefsel dat rijk is aan collageen en elastische vezels, bundels gladde myocyten en vormen het musculoskeletale apparaat van de milt. In het parenchym van de milt worden rode en witte pulp onderscheiden.
Rijst. 25. Lichtmicrofoto van een rattenmilt. Hematoxyline-eosine kleuring. Kwantitatief overheersende rode pulp op het felroze preparaat (oxyfiel), witte pulp - intens basofiel, is een opeenhoping van lymfocyten.
Rijst. 26.
De witte pulp maakt ongeveer 20% van de massa van het orgaan uit en wordt gevormd door lymfocyten, plasmacellen, macrofagen, dendritische en interdigiterende cellen, waarvan het reticulaire weefsel het raamwerk is. Bolvormige ophopingen van dit soort cellen worden lymfatische follikels (knobbeltjes) van de milt genoemd. De diameter van de follikels is 0,3-0,5 mm, ze zijn omgeven door een capsule van reticuloendotheelcellen.
Rijst. 27. Semi-schematische weergave van lichtmicroscopie van een fragment van de milt. Hematoxyline-eosine kleuring.
De lymfatische follikel van de milt (Malpighiaans lichaam) heeft vier zones: peri-arterieel, mantel, marginaal en ook een licht (reactief of germinaal) centrum. De reactieve centra van de lymfatische follikels van de milt en lymfeklier zijn identiek in structuur en functie van de formatie. Ze bevatten B-lymfoblasten, typische macrofagen, dendritische en reticulaire cellen. Het verschijnen van reactieve centra in de follikels is een reactie op antigene stimulatie. De periarteriële zone is een opeenhoping van T-lymfocyten rond de slagader van de lymfatische follikel, of, zoals het ook wordt genoemd, de centrale slagader van de milt. De peri-arteriële zone is verrijkt met interdigiterende cellen - macrofagen, die in staat zijn om complexen van antilichamen met antigenen op hun oppervlak te fixeren en de proliferatie en rijping van T-lymfocyten te veroorzaken. De peri-arteriële zone van de miltfollikels is een analoog van de thymus-afhankelijke paracorticale zone van de lymfeklieren. De donkere mantelzone wordt gevormd door compact gelegen kleine B-lymfocyten en een klein aantal T-lymfocyten, plasmacellen en macrofagen. De marginale zone - de plaats waar de witte pulp overgaat in de rode - wordt gevormd door B- en T-lymfocyten, macrofagen en wordt begrensd door poreuze sinusoïdale hemocapillairen. Na de rijping van lymfocyten gaan ze van het lichte centrum en de peri-arteriële zone naar de mantel en de marginale zones met de volgende uitgang in de bloedbaan.
Rijst. 28. Lichtmicrofoto van een fragment van het lichaam van Malpighian, x 400. Gekleurd met hematoxyline-eosine. In het midden van de follikel is een licht kiemcentrum zichtbaar, aan de periferie waarvan zich een centrale slagader bevindt.
De marginale zone, als gevolg van contacten met hemocapillairen, accumuleert een grote hoeveelheid antigenen uit het bloed en speelt daarom een belangrijke rol bij de immunologische activiteit van de milt. Een groot aantal pulpa-arteriolen die uit de centrale slagader komen, verlaten de witte pulp, maar keren dan terug en stromen in de sinussen van de marginale zone rond de knoop. Een groot aantal macrofagen en reticulo-endotheelcellen langs de periferie van de follikel dienen voor fagocytose en verwijdering van antigeen afval uit het bloed. Dendritische cellen die zich hier bevinden, absorberen en verzenden antigenen naar immunologisch competente cellen (T- en B-lymfocyten), die de systemische circulatie in de witte pulp verlaten, precies vanuit de sinusoïdale haarvaten van de marginale zone van de follikel. Geactiveerde lymfocyten migreren naar het kiemcentrum van de knobbel, veranderen in immunoblasten (de zogenaamde blasttransformatie van lymfocyten vindt plaats), prolifereren en veranderen in effectorcellen. Deze laatste komen de rode pulp binnen, waar de plasmacellen clusters vormen in de vorm van Billrott-strengen en antilichamen produceren die in het bloed worden afgegeven. Geactiveerde T-lymfocyten verlaten de rode pulpa en keren terug naar de algemene bloedsomloop.
Rijst. 29. Thymus-afhankelijke en thymus-onafhankelijke zones van de milt. De opeenhoping van T-lymfocyten (lichtcellen) rond de slagaders die uit de trabeculae komen, vormt een thymusafhankelijke zone. De lymfatische follikel en het lymfoïde weefsel van de witte pulp eromheen is een thymus-onafhankelijke zone. Hier zijn B-lymfocyten (donkere cellen), macrofagen en folliculaire procescellen aanwezig.
Lymfatische peri-arteriële omhulsels zijn een langwerpige opeenhoping van lymfocyten, die in de vorm van klauwen de slagaders van de witte pulp bedekken en aan de ene kant doorgaan in de lymfatische follikels van de milt. In het centrale deel van de vagina, dichter bij het lumen van het vat, zijn B-lymfocyten en plasmocyten geconcentreerd, aan de periferie - T-lymfocyten.
De rode pulp, die ongeveer 80% van de massa van de milt uitmaakt, is een opeenhoping van bloedcellen die zich ofwel in de omgeving van de reticulaire cellen of in het systeem van de vasculaire sinussen van de milt bevinden. Gebieden met rode pulp die zich tussen de sinussen bevinden, worden pulpakoorden van de milt genoemd. Ze voeren de processen uit van transformatie van B-lymfocyten in plasmacellen, evenals monocyten in macrofagen. Miltmacrofagen zijn in staat om oude of beschadigde rode bloedcellen en bloedplaatjes te herkennen en te vernietigen. In dit geval wordt het hemoglobine van de vernietigde erytrocyten gebruikt en wordt het een bron van ijzer voor de synthese van bilirubine en transferrine. De moleculen van de laatste worden door macrofagen van het rode beenmerg aan de bloedsomloop onttrokken en gebruikt bij het proces van erytrocytneogenese.
Rijst. dertig. Lichtmicrofoto van een fragment van de rode pulp van de milt, x 1000. Gekleurd met hematoxyline-eosine. Talrijke macrofagen worden gezien omringd door bloedcellen. Bleek gekleurde procesvormige cellen - stromale elementen van de milt - reticulaire cellen.
Rijst. 31. Elektronenmicrofoto van een fragment van de rode pulp van de milt. Men kan de poreuze endotheelwand van de veneuze sinussen en de lymfoïde banden van Billrott daartussen zien.
Rijst. 32. Scanning elektronenmicroscopie. Fragment van de veneuze sinus van de milt. Je kunt de opening zien tussen de endotheliocyten, waarin de gevormde elementen van het bloed worden geperst. De letters geven aan: N - neutrofiel; M - macrofaag, L - lymfocyt. Macrofagen regelen de kwaliteit van cellen die terugkeren naar de bloedbaan.
Het vasculaire systeem van de milt heeft een aantal kenmerken die ervoor zorgen dat de functies van dit orgaan worden uitgevoerd. Dientengevolge komt de miltslagader de poorten van de milt binnen, die vertakt in een systeem van vertakkingen in de trabeculae van de milt, de trabeculaire slagaders. De trabeculaire slagaders verdelen zich in de slagaders van de witte pulp van de milt, waarrond lymfocyten zijn gegroepeerd en peri-arteriële lymfatische omhulsels en follikels van de milt worden gevormd. Die delen van de witte pulpaslagaders die door de lymfatische follikels gaan, worden centrale slagaders genoemd, omdat ze dienen als centra voor de uitdrijving van lymfocyten tijdens de vorming van lymfatische follikels in ontogenese. De centrale slagaders gaan over in de slagaders van de rode pulpa, deze vallen uiteen in borstelarteriolen, die eindigen in ellipsoïde (mouw)arteriolen. Ellipsoïde arteriolen zijn omgeven door een soort "mouwen" - koppelingen van clusters van reticulaire cellen en reticulaire vezels die de rol spelen van arteriële sluitspieren van de milt. Via een systeem van hemocapillairen communiceren ellipsoïde arteriolen met de veneuze sinussen van de milt van een poreus type. Dit is het zogenaamde systeem van gesloten (gesloten) bloedcirculatie van de milt. Sommige van de haarvaten kunnen echter direct in de rode pulp openen en een open (open) miltcirculatiesysteem vormen. Veneuze sinussen met een aanzienlijke bloedvulling kunnen dienen als bloeddepot. Vanuit de veneuze sinussen stroomt bloed in de aderen van de rode pulp, vervolgens in de trabeculaire aderen en van de laatste in de miltader. In de wand van de veneuze sinussen in het gebied van hun overgang nabij de ader van de rode pulp, is er een opeenhoping van gladde myocyten die de veneuze sluitspieren van de milt vormen.
Rijst. 33. Schema van bloedtoevoer naar de milt Trabeculaire slagaders → pulpslagaders → arteriolen en haarvaten van de follikel → sinussen van de marginale zone → uitgang van T- en B-lymfocyten uit het vaatbed. Follikelarteriolen → borstelarteriolen van de rode pulp → sinusoïde haarvaten.
Rijst. 34. Sinusoïden in de rode pulp van de milt. Volgens de theorie van open circulatie (hierboven) komt bloed uit de haarvaten de rode pulp binnen en vervolgens in de sinusoïden. Volgens de theorie van de gesloten circulatie (van onderaf) openen de haarvaten direct in sinusoïden.
Rijst. 35. Lichtmicroscopie van miltfragmenten met verschillende histologische kleuringen: A - impregnatie met zilvernitraat (reticulair stroma is zichtbaar); B - kleuring met hematoxyline-eosine (structurele componenten van het orgelparenchym worden gevisualiseerd); C – kleuring met ijzerhematoxyline (verschillende dichtheid van lymfoïde elementen in de witte pulp is duidelijk zichtbaar).
Met de samentrekking van de veneuze sluitspieren hoopt het bloed zich op in de sinussen, het wordt dikker als gevolg van de impregnatie van het plasma door de wand van de veneuze sinussen. Bij gelijktijdige samentrekking van arteriële en veneuze sluitspieren wordt bloed in de milt afgezet. Ontspanning van arteriële en veneuze sluitspieren met gelijktijdige samentrekking van gladde myocyten van het kapsel en trabeculae van de milt bepaalt vooraf de afgifte van afgezet bloed in het veneuze bed.
Het leggen van de milt wordt uitgevoerd aan het begin van de tweede maand van de embryonale ontwikkeling in de vorm van ophopingen van mesenchymale cellen doordrongen van bloedvaten in het dorsale mesenterium. Uit het mesenchym wordt reticulair weefsel gevormd, dat laatste wordt bevolkt door bloedstamcellen. In de derde maand van embryogenese differentieert de peri-arteriële thymus-afhankelijke zone in de milt, in de vijfde maand worden reactieve centra en marginale zones van follikels gevormd, in de zesde maand kan rode pulp worden onderscheiden. Tegelijkertijd (van de derde tot de vijfde maand van de embryogenese) nemen de verschijnselen van myeloïde hematopoëse toe in de milt, het vervult de functies van een universeel hematopoëtisch orgaan. Vanaf de zesde maand tot de geboorte van een kind vervagen de manifestaties van myeloïde hematopoëse, ze worden vervangen door de processen van lymfocytopoëse. Op volwassen leeftijd vertoont de milt aanzienlijke herstellende vermogens; experimenteel bewezen de mogelijkheid van vernieuwing met het verlies van 80-90% van het parenchym. De massa van de milt neemt iets af op de leeftijd van 20 tot 30 jaar; in het interval van 30 tot 60 jaar is het stabiel. Op oudere leeftijd werden atrofie van de rode en witte pulp, proliferatie van bindweefselstroma, een afname van het gehalte aan macrofagen en lymfocyten onder parenchymale elementen, een toename van het gehalte aan granulocyten en weefselbasofielen en het verschijnen van megakaryocyten opgemerkt. Het gebruik van ijzer uit in de milt vernietigde erytrocyten gaat achteruit.
Intercellulaire interacties
bij het leveren van de immuunafweer van het lichaam.
Voor een adequate reactie op lichaamsvreemde stoffen (antigene stimulatie) is interactie en samenwerking van verschillende typen cellen van het immuunsysteem noodzakelijk. Onder hen worden cellen van macrofagen onderscheiden - monocyten bloed, histiocyten- bindweefselmacrofagen, beenmerg, peritoneale, alveolaire macrofagen, Langerhans-cellen huid, Kashchenko-Hofbauer-cellen placenta, stellate reticuloendotheliocyten lever, dendritisch en interdigiterende cellen lymfeklieren en milt, osteoclasten botweefsel microgliocyten zenuwstelsel. Er is een groep zogenaamde microfagen, waartoe ze behoren neutrofiele granulocyten bloed, evenals cellen die, onder bepaalde omstandigheden van functioneren, fagocytische eigenschappen kunnen vertonen, - endotheliocyten, fibroclasten. Ten slotte combineert de derde groep cellen verschillende populaties van T- en B-lymfocyten ( T-killers, T-helpers, T-suppressors, plasmacellen, T- en B-geheugencellen). De totale massa cellen die direct zorgen voor de immuunafweer van het lichaam is ongeveer 1% van het lichaamsgewicht.
Allereerst reageren T-helpers op het binnendringen van vreemde deeltjes in het lichaam: antigene determinanten binden aan specifieke receptoren op hun oppervlak. Het gevormde antigeenreceptorcomplex komt los van het oppervlak van het T-helperplasmolema en wordt gefixeerd door de oppervlaktereceptoren van de macrofaag. In het volgende stadium worden door macrofagen gemodificeerde antigenen overgebracht naar B-lymfocyten, die, onder invloed van antigene stimulatie en de activerende werking van T-helpers, in plasmacellen veranderen. Deze laatste synthetiseren eiwitmoleculen van immunoglobulinen (antilichamen), die selectief binden aan antigenen en hun inactivatie vooraf bepalen. T-helpers produceren, na contact met het antigeen, speciale chemicaliën die de proliferatie van T-killers stimuleren. Deze laatste hebben het vermogen om de celmembranen van bacteriën en cellen die antigene determinanten op hun oppervlak dragen, te vernietigen.
In elk van deze stadia kan gedeeltelijke inactivering van vreemd materiaal optreden, evenals de modificatie en overdracht naar andere celpopulaties om een immuunrespons te ontwikkelen. Het is mogelijk dat een antigeenbevattend deeltje wordt herkend en afgevoerd door een macrofaag zonder de deelname van een T-lymfocyt, wordt gesplitst door zijn lysosomale enzymen, en de resulterende antigene fragmenten worden overgebracht naar T- en B-lymfocyten en stimuleren hun transformatie in effectorcellen (T-killers en plasmacellen), evenals geheugencellen.
Interactie van cellen in de immuunrespons
De immuunrespons is mogelijk als gevolg van de activering van lymfocytklonen en bestaat uit twee fasen. In de eerste fase activeert het antigeen die lymfocyten die het herkennen. In de tweede (effector)fase coördineren deze lymfocyten de immuunrespons gericht op het elimineren van het antigeen.
humoristisch immuun antwoord
De humorale immuunrespons omvat macrofagen (antigeenpresenterende cellen), T-helpers en B-lymfocyten.
macrofaag absorbeert het antigeen dat het lichaam binnendringt en onderwerpt het aan verwerking - splitsing in fragmenten. Antigeenfragmenten worden samen met het MHC-molecuul op het celoppervlak blootgesteld. Het complexe "antigeenmolecuul MHC klasse II" wordt aangeboden aan T-helper (Fig. 35).
Rijst. 36. Antigeenherkenning door T-lymfocytreceptor. Met behulp van de T-lymfocytreceptor herkent de T-cel het antigeen, maar alleen in complex met het MHC-molecuul. In het geval van de T-helper is zijn molecuul, CD4, betrokken bij het proces, dat zich met zijn vrije uiteinde aan het MHC-molecuul bindt. Het antigeen dat door de T-cel wordt herkend, heeft twee plaatsen: de ene interageert met het MHC-molecuul, de andere (epitoop) bindt aan de T-lymfocytreceptor. Een soortgelijk type interactie, maar met de deelname van het CD8-molecuul, is kenmerkend voor het proces van herkenning door de T-killer van het antigeen dat is geassocieerd met het MHC klasse I-molecuul.
T-helper herkent het antigeen-MHC klasse II-molecuulcomplex op het oppervlak van de antigeenpresenterende cel. Het herkenningsproces omvat de interactie van het T-celreceptor-CD3-complex, dat zorgt voor specificiteit en deelname van ondersteunende co-stimulerende moleculen. Om de T-helper te activeren, is specifieke herkenning door de T-helper van een antigeenfragment op het oppervlak van een antigeenpresenterende cel onvoldoende. Activering van T-helpers wordt verschaft door de interactie van het B7-molecuul (CD80) op het oppervlak van de antigeenpresenterende cel met het CD28-molecuul op het oppervlak van de T-helper. T-helpers zijn bijzonder gevoelig voor stimulatie via CD28, die differentiëren tot T-helper 2 en B-cellen activeren via CD80. Bij zwakke expressie van CD28 en in aanwezigheid van het CTLA-molecuul ( C ytotoxischT - ik ymfocytp rotin) T-helpers 1 worden gevormd.
Herkenning door T-helper van de gewenste moleculen op het oppervlak van de antigeenpresenterende cel stimuleert de secretie van IL1. Geactiveerde IL1 T-helper synthetiseert IL2- en IL2-receptoren, waardoor de agonist de proliferatie van T-helpers en cytotoxische T-lymfocyten stimuleert. In het geval van T-helper hebben we het over autocriene stimulatie, wanneer de cel reageert op het middel dat zelf synthetiseert en afscheidt. Dus, na interactie met een antigeenpresenterende cel, verwerft T-helper het vermogen om te reageren op de werking van IL2 met een uitbarsting van proliferatie. De biologische betekenis van dit proces is de accumulatie van een dergelijke hoeveelheid T-helpers, die zal zorgen voor de vorming in de lymfoïde organen van het vereiste aantal plasmacellen die in staat zijn antilichamen tegen dit antigeen te produceren.
B-lymfocyt. B-lymfocytactivering omvat directe interactie van het antigeen met een immunoglobuline (Ig) op het oppervlak van de B-cel. In dit geval verwerkt de B-lymfocyt zelf het antigeen en presenteert zijn fragment in verband met het MHC II-molecuul op zijn oppervlak. Dit complex herkent een T-helper geselecteerd door hetzelfde antigeen dat betrokken was bij de selectie van deze B-lymfocyt. Twee paar moleculen zijn betrokken bij de activering van B-cellen: enerzijds de specifieke interactie van het antigeen met de Ig M-receptor op het oppervlak van de B-lymfocyt en anderzijds het CD40-molecuul op het oppervlak van de B-cel interageert met het CD40L (CD154) molecuul op het oppervlak van de T-helper, waardoor de B-cel wordt geactiveerd. Herkenning door de T-helperreceptor van het “antigen-molecuul MHC klasse II”-complex op het oppervlak van de B-lymfocyt leidt tot de secretie van IL2, IL4, IL5 en γ-IFN uit de T-helper, onder invloed van waarbij de B-cel wordt geactiveerd en prolifereert, waardoor een kloon wordt gevormd. De geactiveerde B-lymfocyt differentieert tot een plasmacel: het aantal ribosomen neemt toe, het granulaire endoplasmatisch reticulum en het Golgi-complex worden meer uitgesproken.
Plasma cel synthetiseert Ig. IL6 uitgescheiden door geactiveerde T-helpers stimuleert de secretie van Ig. Een deel van rijpe B-lymfocyten circuleert na antigeenafhankelijke differentiatie in het lichaam als geheugencellen.
mobiel immuun antwoord
De cellulaire immuunrespons wordt gekenmerkt door de proliferatie van toegewijde immunocompetente cellen die reageren met het antigeen in combinatie met het molecuul MHC klas l op het oppervlak van vreemde cellen of endogene antigenen in combinatie met het MHC klasse I-molecuul op het oppervlak van de eigen met virus geïnfecteerde en getransformeerde (tumor)cellen. De cytotoxische T-lymfocyt is betrokken bij de cellulaire immuunrespons.
reacties celgemedieerd cytolyse. Effectorcellen gebruiken hun receptoren om de doelcel te herkennen en te vernietigen. Niet alleen T-lymfocyten, maar ook andere subpopulaties van lymfoïde cellen en, in sommige gevallen, myeloïde cellen zijn verantwoordelijk voor celgemedieerde cytolyse. Het herkenningsproces omvat verschillende moleculen die worden blootgesteld aan het oppervlak van interagerende cellulaire partners:
√ specifieke antigenen, bijvoorbeeld virale peptiden op het oppervlak van geïnfecteerde cellen, in combinatie met het MHC-molecuul, worden herkend door cytotoxische T-celreceptoren, voornamelijk CD8+- en sommige subpopulaties van CD4+-cellen;
√ antigene determinanten van tumorcellen worden herkend door NK-cellen zonder deelname van MHC klasse I-molecuul;
√ antigeen-geassocieerde AT op het oppervlak van doelcellen wordt herkend door receptoren van Fc-fragmenten van NK-cellen (fenomeen van AT-afhankelijke cytotoxiciteit).
cytotoxisch T-lymfocyt(TC). Het antigeen dat op het oppervlak van de doelcel wordt gepresenteerd in een complex met het klasse I MHC-molecuul, bindt aan de cytotoxische T-lymfocytreceptor. Bij dit proces is het CD8-molecuul van het celmembraan Tc betrokken. IL2 uitgescheiden door T-helpers stimuleert de proliferatie van cytotoxische T-lymfocyten.
Verwoesting cellen-doelen. De cytotoxische T-lymfocyt herkent de doelcel en hecht eraan. Kleine korrels zijn aanwezig in het cytoplasma van een geactiveerde cytotoxische T-lymfocyt. Ze bevatten het cytolytische eiwit perforine. De perforinemoleculen die vrijkomen door de T-killer worden gepolymeriseerd in het membraan van de doelcel in aanwezigheid van Ca2+. De perforineporiën gevormd in het plasmamembraan van de doelcel laten water en zouten door, maar geen eiwitmoleculen. Als polymerisatie van perforine plaatsvindt in de extracellulaire ruimte of in het bloed, waar een overmaat aan calcium aanwezig is, dan zal het polymeer niet in staat zijn het membraan binnen te dringen en de cel te doden. De T-killer zelf is beschermd tegen het cytotoxische effect van perforine.
In praktisch werk worden de volgende histologische preparaten voor studie aangeboden:
1. SMEER VAN ROOD BEENMERG.
Kleuring: volgens Romanovsky-Giemsa (azuur II, eosine).
Vind bij lage en dan bij hoge vergroting van de microscoop sinusoïdale capillairen in het preparaat. In hun lumen zijn erytrocyten en leukocyten zichtbaar. Tussen de sinusoïdale haarvaten bevindt zich een reticulair weefsel, in de lussen waarvan hematopoëtische cellen zich in verschillende stadia van rijping bevinden. Cellen van de erytropoëtische reeks: a) proerytroblasten - grote cellen (15 micron) met een grote ronde kern, waarin de nucleoli duidelijk zichtbaar zijn. Een sterk basofiel cytoplasma omringt de kern met een smalle strook; b) basofiele erytroblasten - cellen (10-12 micron) met basofiel cytoplasma. De kern is rond, bevat meer heterochromatine en heeft een donkere kleur; c) polychromatofiele erytroblasten - zelfs kleinere cellen (8-10 micron), hun cytoplasma wordt gelijktijdig gekleurd met zowel zure als alkalische kleurstoffen en ziet er grijsachtig groenachtig uit. De kernen zijn intens gekleurd en hebben geen nucleoli; d) oxyfiele erytroblasten (normoblasten) - het cytoplasma van cellen bevat al een aanzienlijke hoeveelheid hemoglobine, daarom kleurt het oxyfiel, de kernen worden gepycnotiseerd, vaak gelyseerd en verdwijnen; e) reticulocyten - jonge erytrocyten. In het cytoplasma van deze cellen kan men granulaire mesh-structuren zien - de overblijfselen van de kern en het organel; g) rijpe erytrocyten - niet-nucleaire bloedcellen, met een homogeen oxyfiel cytoplasma.
Cellen van de granulocytopoëtische reeks: a) promyelocyten - grote cellen met grote ronde en lichte kernen. Het cytoplasma is matig basofiel, het bevat enkele azurofiele korrels (lysosomen); b) myelocyten bevatten specifieke granulariteit in het cytoplasma: eosinofiele hebben grote helderrode eosinofiele korrels; basofiel - donkerblauwe basofiele korrels; neutrofiel - hebben twee soorten kleine korrels die zowel basische als zure kleurstoffen waarnemen. Myelocyten rijpen tot metamyelocyten. Een kenmerkend morfologisch kenmerk van deze overgangsvorm van cellen is een afname van het volume van het cytoplasma en de kern, die de vorm aannemen van een gebogen staaf of hoefijzer. Afhankelijk van de korrels in het cytoplasma, worden metamyelocyten ook verdeeld in basofiel, oxyfiel en neutrofiel. In de loop van de rijping worden de kernen van metamyelocyten gesegmenteerd en veranderen de cellen in rijpe gesegmenteerde eosinofielen, basofielen en neutrofielen.
In een uitstrijkje van rood beenmerg zijn megakaryocyten gelokaliseerd in de buurt van de sinusoïden - zeer grote (> 50 m) cellen met verschillende polyploïde kernen en basofiel cytoplasma.
Onderzoek het monster, teken en label: 1. Proerytroblasten. 2. Polychromatofiele erytroblasten. 3. Neutrofiele metamyelocyten. 4. Megakaryocyten. 5. Rijpe bloedcellen (erytrocyten, leukocyten) in de sinusoïdale hemocapillair.
Door welke morfologische kenmerken kan een polychromatofiele proerytrocyt worden onderscheiden van een erytrocyt?
In welk stadium van myelocytdifferentiatie kunnen basofiele, oxyfiele en neutrofiele cellen worden geïdentificeerd?
Welke cel is in een rood beenmerguitstrijkje groter: een megakaryoblast of een megakaryocyt?
Benoem op basis van de studie van een uitstrijkje van rood beenmerg de cellen die qua grootte overeenkomen met erytrocyten.
In welk stadium van differentiatie verliest de myelocyt zijn vermogen om te delen?
2. GOIMER (Thymus).
Bij lage vergroting is de lobvormige structuur van de klier zichtbaar. Elke lobule heeft een donkerder perifeer deel - de corticale substantie en een lichter binnenste deel - de medulla. Bij hoge vergroting, in het middelste deel van de medulla, zijn de epitheellichamen van Hassall zichtbaar - een concentrische gelaagdheid van epithelioreticulocyten. De stromale elementen van de lobulus zijn epithelioreticulocyten, die in contact komen met hun processen en een netwerk vormen. In de intervallen tussen de epithelioreticulocyten bevinden zich parenchymale elementen: T-lymfocyten (thymocyten) en macrofagen.
Onderzoek het monster, teken en label: 1. Klierlobule: 1. Cortex: a) T-lymfocyten; 2. Beenmerg: b) epithelioreticulocyten; c) Hassalls lichamen. II. Interlobulair bindweefsel. 3. Bloedvaten.
Maak op basis van het onderzoek van het thymuspreparaat een conclusie over de intensiteit van de mitosen in het subcapsulaire gebied, de cortex en de medulla.
Wat zijn de lichamen van Hassall en waar zijn ze gelokaliseerd in de thymus?
Is het epitheel dat het stroma van de thymus vormt eenlagig of meerlagig? Leg uit waarom.
3. LYMFE KNOOP.
Met een kleine vergroting van de microscoop is te zien dat de lymfeklier is bedekt met een bindweefselcapsule, van waaruit dunne partities, trabeculae, zich naar binnen uitstrekken. Tussen de trabeculae bevindt zich reticulair weefsel dat is geïnfiltreerd door talrijke lymfocyten. Lymfocyten zijn geconcentreerd langs de periferie van de knoop in de vorm van grote clusters van afgeronde vorm - follikels, die de corticale substantie van de lymfeknoop vormen. Van de follikels naar de diepte van de knoop vertrekken de hersenkoorden die de medulla vormen. Lichte ruimtes gevuld met reticulair weefsel en een klein aantal lymfocyten zijn sinussen. De marginale sinus, gelegen tussen de follikels en de capsule, gaat over in de intermediaire corticale sinussen, die op hun beurt doorgaan in de tussenliggende cerebrale sinussen, die lymfe verzamelen in de centrale sinus bij de poort van de lymfeklier.
Maak een schets van het preparaat en benoem: 1. Capsule. 2. Trabeculae. 3. Cortex. 4. De medulla. 5. Follikels. 6. Hersenbanden. 7. Marginale sinus. 8. Tussenliggende corticale sinussen. 9. Tussenliggende cerebrale sinussen. 10. Poort van de lymfeklier. 11. Reticulair weefsel.
Trek op basis van het bestudeerde preparaat een conclusie over de antigene stimulatie van de lymfeklier en licht je antwoord toe.
Wat circuleert er in de sinussen van de lymfeklier?
Geef op het preparaat de plaats aan waar zich de meeste plasmacellen bevinden.
4. MILT.
Gekleurd met hematoxyline en eosine.
Bij een lage vergroting van de microscoop is duidelijk een dicht bindweefselkapsel te zien, van waaruit de trabeculae zich uitstrekken tot in het orgaan. Het kapsel van de milt is bedekt met mesothelium en bevat een kleine hoeveelheid gladde spiercellen, die, door samen te trekken, bijdragen aan de afgifte van bloed door de poorten van het orgel. Tussen de trabeculae bevindt zich de witte en rode pulp van de milt. De witte pulp bestaat uit lymfatische follikels die een centrale slagader hebben. De rode pulp wordt gevormd door reticulair weefsel, bloedcellen, voornamelijk erytrocyten en veneuze sinussen.
Onderzoek het preparaat met een sterke vergroting, teken een klein gebied en wijs aan: 1. Miltcapsule. 2. Trabeculae. 3. Witte pulp (lymfatische follikels): a) centrale slagader; b) kiemcentrum. 4. Rode pulpa: a) reticulair weefsel, b) erytrocyten; c) leukocyten. 5. Veneuze sinus.
Wat zijn de belangrijkste onderscheidende kenmerken van de structuur van de lymfatische follikels van de milt en lymfeklieren.
Maak een conclusie waar er meer lymfoïde knobbeltjes zijn met voortplantingscentra in de milt of lymfeklier en leg uit waarom?
DEMOPRODUCTEN:
1. Een uitstrijkje van het rode beenmerg van een 5-jarig kind.
2. Thymusklier van een 12-jarig kind.
3. Reticulaire vezels in de lymfeklier. Zilver impregnatie.
ELEKTRONISCHE MICROFOTOGRAFIE:
1. Rode beenmergcellen.
2. Epithelioreticulocyten van de thymus.
3. T-lymfocyten.
4. Sinus van de lymfeklier.
5. Sinus van de milt.
Situationele taken:
1. Bij de voorbereiding is er een snee in het buisvormige bot van een kind van 3-5 jaar oud, een jonge man van 12-18 jaar oud en een oud persoon. Hoe verandert de toestand en topografie van het rode beenmerg met de leeftijd?
2. De thymus is verwijderd bij een pasgeboren kind. Als gevolg van deze operatie nam haar vermogen om antistoffen aan te maken sterk af. Leg de reden voor dit fenomeen uit.
3. Er zijn microfoto's van de lymfeklieren van het buikvlies, gefotografeerd op het hoogtepunt van de spijsvertering en in rust. Hoe kan men tijdens de spijsvertering een lymfeklier onderscheiden en dit fenomeen verklaren?
4. Het dier werd direct na de geboorte onder steriele omstandigheden geplaatst. Kunnen zich in deze situatie secundaire follikels in de lymfeklieren vormen, zo ja, waarom, zo niet, waarom?
5. In de oudheid werd bij marathonlopers de milt verwijderd. Leg uit waarom?
LYMPHOID WEEFSEL (anat. lymfa, van lat. lymfa zuiver water, vocht + Grieks -eidēs vergelijkbaar) - is een verzameling lymfocyten en macrofagen die zich in het cellulaire fibreuze reticulaire stroma bevinden. Lymfoïde weefsel vormt het actieve parenchym van de lymfoïde organen. Lymfoïde organen zijn organen van immunogenese, waaronder de thymusklier, lymfeklieren, milt, lymfoïde elementen van het beenmerg en ophopingen van lymfoïde weefsel in de wanden van de luchtwegen, de urinewegen en het maagdarmkanaal.
De basis van lymfoïde weefsel zijn reticulaire vezels en cellen die een netwerk vormen met cellen van verschillende groottes. Dit netwerk is uitgerust met lussen waarin zich lymfoïde cellen, macrofagen, mestcellen en een klein aantal leukocyten bevinden. De vorming van het reticulaire stroma vindt plaats vanuit het mesenchym en de vorming van lymfoïde cellen uit de stamcellen van het beenmerg.
De cellen van de lymfoïde reeks zijn verdeeld in twee groepen::
- T-lymfocyten
- B-lymfocyten.
De beweging van cellen vindt plaats met bloed en lymfe. De functie van cellen van de lymfoïde reeks is om deel te nemen aan de reacties van de immuunrespons gericht tegen genetisch vreemde stoffen.
De structuur van lymfoïde weefsel, de topografie van de elementen van de structuur in verschillende organen van het immuunsysteem onderscheidt zich door zijn eigen kenmerken. Lymfoïde weefsel in de centrale organen van immunogenese handhaaft een functionele eenheid met andere weefsels. In de thymus bijvoorbeeld is lymfoïde weefsel in functionele eenheid met epitheelweefsel en in het beenmerg met myeloïde weefsel. De aanwezigheid van lymfoïde weefsel in verschillende kwalitatieve toestanden hangt rechtstreeks af van de volwassenheid en functionele toestand van de perifere organen van het immuunsysteem, aangezien de aanwezigheid van enkele lymfocyten en lymfoïde knobbeltjes wijst op een verhoogde immuunactiviteit van het lichaam.
Het grootste aantal lymfoïde knobbeltjes kan worden gevonden in organen zoals: amandelen, lymfoïde plaques, milt, wanden van de appendix, maag, dunne en dikke darm, lymfeklieren bij kleine kinderen en adolescenten.
Lymfoïde weefsel vormt een zeldzame en dunne beschermende laag van lymfoïde cellen en bevindt zich onder de epitheliale bedekking van de urinewegen en luchtwegen, maagdarmkanaal. Lymfoïde weefsel draagt bij aan de vorming van lymfoïde moffen rond arteriële vaten in de milt. De afname van de hoeveelheid lymfoïde weefsel en lymfoïde knobbeltjes hangt af van het verouderingsproces van het lichaam. Als ontstekingsprocessen en activering van immuunreacties van primaire en secundaire aard in het lichaam worden waargenomen, treedt daarom reactieve hyperplasie van de lymfeklieren op. De nederlaag van lymfoïde weefsel vindt plaats met histiocyten, paraproteïnemische hemoblastosen, lymfogranulomatose, histiocyten X, kwaadaardige lymfomen.
Levenscyclus van immunocompetente cellen lymfoïde weefsel (B- en T-cellen en hun nakomelingen in de vorm van cellen die immunoglobulinen en antigeenherkennende en effectorlymfocyten synthetiseren) is aanzienlijk korter dan de levensduur van het organisme. In dit opzicht is de belangrijkste voorwaarde voor de implementatie van immunologische reacties de constante stroom van histogenese in het lymfoïde weefsel, waarbij de voorlopers van immunocompetente cellen prolifereren en differentiëren tot rijpe cellen.
De laatste zijn meestal zeer gespecialiseerd en heterogeen en differentiatieverschillen die verband houden met immunologische functies zijn daarin rigide vastgelegd (de klasse en specificiteit van het gesynthetiseerde immunoglobuline in B-cellen en de specificiteit van receptoren voor antigenen in T-cellen). Integendeel, onder de voorlopercellen gedurende het hele leven zijn er ook die waarvoor verschillende differentiatiemogelijkheden openstaan.
Het orgaan van immuniteit - - heeft er nog een belangrijk kenmerk. De meeste van zijn voorlopercellen zijn mobiele, recirculerende elementen. Ze worden gevormd op plaatsen die ver verwijderd zijn van die waar ze vervolgens migreren en waar ze het nageslacht vormen van rijpe cellen.
Dus, processen, die in veel andere weefsels alleen worden uitgevoerd op het moment van leggen tijdens de morfogenese, in het lymfoïde weefsel gedurende het hele leven. Ze zorgen voor een constante verandering in de cellulaire samenstelling, een reeks gesynthetiseerde immunoglobulinen (antilichamen) en soorten immunologisch actieve lymfocyten in overeenstemming met het veranderende spectrum van antigenen die het lymfoïde weefsel binnenkomen.
op lymfoïde weefsel slaagde erin om overtuigend aan te tonen dat deze veranderingen voornamelijk of zelfs uitsluitend worden uitgevoerd door de selectie van toegewijde voorlopercellen, vooraf bepaald voor latere differentiatie, ongeacht de impact van antigenen. Bij de differentiatie van lymfoïde cellen in een volwassen organisme worden dus van nature twee stadia onderscheiden: de ontwikkeling van antigeenonafhankelijke voorlopers en de antigeenafhankelijke differentiatie van hun nakomelingen.
De eerste fase begint met een algemene hematopoietische stamcel en eindigt met toegewijde voorgangers. Het is echter niet bekend in welke volgorde de scheiding in T- en B-precursoren in dit stadium plaatsvindt, in precursors die verschillen in immunologische specificiteit en klassen van gesynthetiseerde immunoglobulinen, evenals in behorend tot verschillende subpopulaties van T-cellen. De resultaten die werden verkregen met het gebruik van transplantatie van beenmergcellen die waren gemarkeerd met chromosomale herschikkingen na bestraling, toonden met name aan dat het beenmerg stamcellen bevat die in staat zijn zichzelf in stand te houden, maar een beperkte differentiatiekracht hebben, bijvoorbeeld cellen die alleen differentiëren in myeloïde of alleen in T-cellen.
Het is echter niet gevonden terwijl stamcellen, verstoken van potentie voor myeloïde differentiatie, maar behield het vermogen om voorlopers te zijn voor zowel T- als B-cellen. Er kan daarom worden aangenomen dat zelfvoorzienende voorlopers van T-cellen (prethymische stamcellen) direct zijn afgeleid van pluripotente hematopoëtische stamcellen zonder het voorloperstadium te doorlopen dat alle lymfoïde cellen gemeen hebben. Wat betreft B-cellen, tussen hen en hematopoëtische stamcellen zijn er mogelijk helemaal geen voorlopers die in staat zijn zichzelf in stand te houden.
De kwestie van de mechanismen die werken onder opleiding van toegewijde voorlopers en het reguleren van de expressie van genen die verantwoordelijk zijn voor immunologische functies daarin, blijft open. Er zijn goede redenen om aan te nemen dat deze belangrijke fase in de ontwikkeling van immunocompetente cellen antigeenonafhankelijk is. Integendeel, de daaropvolgende ontwikkeling van reeds geëngageerde cellen en hun nakomelingen is duidelijk antigeenafhankelijk. Het zorgt voor een verandering in de cellulaire samenstelling van het lymfoïde weefsel door voornamelijk die voorlopers te vermenigvuldigen en te differentiëren waarvan de oppervlaktereceptoren de antigenen die er op dit moment in aanwezig zijn, herkennen.
Een ander mechanisme voor de selectieve proliferatie van toegewijde voorgangers geassocieerd met cellulaire interacties waarbij een door antigeen geactiveerde cel aangrenzende progenitorcellen stimuleert. In beide gevallen werken antigenen als initiatoren van veranderingen in de samenstelling van celpopulaties die het lymfoïde weefsel bewonen, en als inductoren van eiwitten die door dit weefsel worden gesynthetiseerd.
Lymfoïde weefsel is zeer gevoelig voor externe en interne invloeden. Naarmate het lichaam ouder wordt, neemt de hoeveelheid Lt af. en lymfoïde knobbeltjes in de organen van het immuunsysteem.
Lymfoïde weefsel (synoniem met lymfatisch weefsel) is een verzamelnaam voor de structuren waarin de vorming van lymfocyten plaatsvindt. Menselijk lymfoïde weefsel maakt ongeveer 1% van het lichaamsgewicht uit en is een van de belangrijkste componenten van lymfoïde organen.
Wat is hypertrofie van het lymfoïde weefsel van de keelholte -
Een van de belangrijkste functies van de lymfoïde organen is hun deelname aan de processen van hematopoëse (lymfopoëse). Een belangrijke functie van lymfoïde weefsel is geassocieerd met dit vermogen van lymfocyten - zijn deelname aan de afweerreacties van het lichaam. Grote invloed op de mate van ontwikkeling van lymfoïde weefsel hebben hormonen van de bijnierschors. Onvoldoende functie van de bijnierschors veroorzaakt de groei van lymfoïde weefsel. De introductie van hormonen van de bijnierschors leidt tot degeneratie van lymfoïde weefsel en de dood van lymfocyten.
De structuur en rol van lymfoïde weefsel in de activiteit van het immuunsysteem
De structuur van L.t., de topografie van zijn structurele elementen in verschillende organen van het immuunsysteem heeft zijn eigen kenmerken. In de centrale organen van immunogenese L.t. is in functionele eenheid met andere weefsels, bijvoorbeeld in het beenmerg - met myeloïde weefsel, in de thymus - met epitheelweefsel. Naast ophopingen bevindt L. t. zich in de vorm van een zeldzame, dunne, als het ware beschermende laag van lymfoïde cellen onder de epitheliale bedekking van de luchtwegen en de urinewegen en het maagdarmkanaal.
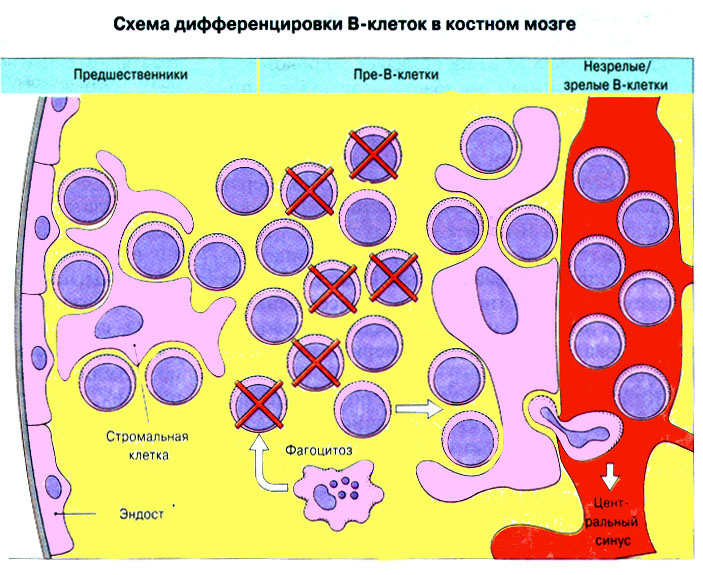
Lymfoïde weefsel van de slijmvliezen: een introductie
Lymfoïde organen worden geclassificeerd als primaire (centrale) of secundaire organen. Lymfocyten behoren dus tot de categorie cellen die wijd verspreid in het lichaam zijn. Lymfoïde weefsel is een type bindweefsel dat wordt gekenmerkt door een hoog gehalte aan lymfocyten.
In de meeste lymfoïde organen vormen fibroblastachtige reticulaire cellen deze vezels, waarop hun talrijke processen zich bevinden. Nodulair lymfoïde weefsel wordt gevormd door bolvormige ophopingen van lymfocyten; dit zijn de zogenaamde lymfoïde knobbeltjes, of lymfoïde follikels, die voornamelijk B-lymfocyten bevatten. Mucosaal-geassocieerd lymfoïde weefsel waarvan de beschermende werking gebaseerd is op de productie van IgA wordt vaak afgekort als MALT (mucosal-associated lymphoid tissue).
De linguale tonsil bestaat uit ophopingen van lymfoïde weefsel - lymfoïde knobbeltjes, waarvan het aantal (80-90) het grootst is in de kindertijd, adolescentie en adolescentie. Tegen de tijd van geboorte neemt het aantal lymfoïde knobbeltjes in de zich ontwikkelende amandel aanzienlijk toe. Reproductiecentra in lymfoïde knobbeltjes verschijnen kort na de geboorte (in de 1e levensmaand). In de toekomst neemt hun aantal toe tot de adolescentie.
De structuur van lymfoïde weefsel. Histologie, functies
Takken van de rechter en linker linguale slagaders, evenals, in zeldzame gevallen, takken van de gezichtsslagader, naderen de linguale amandel. Vanaf deze plaat in de mediale richting strekken trabeculae (septa) zich uit in het lymfoïde weefsel van het orgaan, dat, indien goed uitgesproken, de tonsil in lobben verdeelt.
Bij een 5 maanden oude foetus wordt de amandel weergegeven door een opeenhoping van lymfoïde weefsel tot 2-3 mm groot. Tijdens deze periode beginnen epitheliale strengen te groeien in de vormende amygdala - toekomstige crypten worden gevormd. Op het oppervlak van de plooien bij kinderen zijn talrijke kleine knobbeltjes zichtbaar, in de diepten waarvan zich ophopingen van lymfoïde weefsel bevinden - lymfoïde knobbeltjes.
Onder de epitheliale bedekking in diffuus lymfoïde weefsel bevinden zich lymfoïde knobbeltjes van de faryngeale tonsil met een diameter tot 0,8 mm, waarvan de meeste reproductiecentra hebben. De keelholte tonsil wordt gelegd op de 3-4e maand van het intra-uteriene leven in de dikte van het opkomende slijmvlies van het neusgedeelte van de keelholte.
Tegen het einde van het jaar bereikt de lengte 12 mm en de breedte 6-10 mm. lymfoïde knobbeltjes in de amandel verschijnen in het 1e levensjaar. Na 30 jaar neemt de grootte van de faryngeale tonsil geleidelijk af. De leeftijdsgebonden involutie van de tuba tonsil begint in de adolescentie en adolescentie. Het wordt meestal waargenomen bij kinderen van 3-10 jaar. Hypertrofisch lymfoïde weefsel ondergaat fysiologische involutie en neemt af tijdens de puberteit.
Met behoud van zijn functie kan hypertrofisch lymfoïde weefsel echter pathologische veranderingen in de neus, oren en strottenhoofd veroorzaken. Hypertrofie van de palatinale amandelen wordt vaak gecombineerd met hypertrofie van de gehele faryngeale lymfoïde ring, vooral met hypertrofie van de faryngeale tonsil. Tijdens de puberteit ondergaan de adenoïden regressie, maar de resulterende complicaties blijven en leiden vaak tot invaliditeit. Indirecte tekens adenoïden is ook hypertrofie van de palatine amandelen en lymfoïde elementen op de achterkant van de keelholte.
Hypertrofie van lymfoïde weefsel als reactie op een infectieziekte leidt tot een toename van ontstekingsprocessen in de keelholte. In de dikte van de amandel zijn er ronde dichte ophopingen van lymfoïde weefsel - lymfoïde knobbeltjes van de amandel. Plots van lymfoïde weefsel bevinden zich in het slijmvlies van sommige organen (bronchiën, urinewegen, nieren).
Vocht + Grieks. -eid is vergelijkbaar) een complex van lymfocyten en macrofagen in het cellulaire fibreuze reticulaire stroma; vormt het functionerende parenchym van de lymfoïde organen. De lymfoïde organen, die organen van immunogenese zijn, omvatten de thymusklier (thymusklier) ,
de lymfeklieren ,
milt (milt) ,
lymfoïde elementen van het beenmerg en ophopingen van L.t. in de wanden van het maagdarmkanaal, de luchtwegen en de urinewegen. Basis L.t. vormen reticulaire vezels en reticulaire cellen en vormen een netwerk met cellen van verschillende groottes. De lussen van dit netwerk bevatten cellen van de lymfoïde reeks (kleine, middelgrote en grote lymfocyten, plasmacellen, jonge cellen - blasten), macrofagen, evenals een klein aantal leukocyten, mestcellen. De reticulaire wordt gevormd uit het mesenchym en de cellen van de lymfoïde reeks worden gevormd uit de stamcellen van het beenmerg. Cellen van de lymfoïde reeks, waaronder twee populaties worden onderscheiden - T- en B-lymfocyten, bewegen met bloed en lymfe. Samen met macrofagen zijn ze betrokken bij immuunreacties tegen genetisch vreemde stoffen (zie Immuniteit) .
De structuur van L.t., de topografie van zijn structurele elementen in verschillende organen van het immuunsysteem heeft zijn eigen kenmerken. In de centrale organen van immunogenese L.t. is in functionele eenheid met andere weefsels, bijvoorbeeld in het beenmerg - met myeloïde weefsel, in de thymus - met epitheelweefsel. In de perifere organen van het immuunsysteem, bijvoorbeeld in de wanden van het maagdarmkanaal, de luchtwegen en de urinewegen, afhankelijk van de mate van rijpheid en functionele toestand van L.t. bevindt zich in verschillende kwalitatieve toestanden - van enkele lymfocyten en diffuus gelegen lymfoïde weefsel tot lymfoïde knobbeltjes met reproductiecentra, waarvan de aanwezigheid wijst op een hoge immuunactiviteit van het lichaam. Het grootste aantal lymfoïde knobbeltjes, inclusief die met reproductiecentra, wordt gevonden in de amandelen, lymfoïde plaques, milt, wanden van de appendix, maag, dunne en dikke darm en in de lymfeklieren van kinderen en adolescenten. Naast ophopingen bevindt L. t. zich in de vorm van een zeldzame, dunne, als het ware beschermende laag van lymfoïde cellen onder de epitheliale bedekking van de luchtwegen en de urinewegen en het maagdarmkanaal. In de milt vormt het lymfoïde moffen rond arteriële vaten. Naarmate het lichaam ouder wordt, neemt de hoeveelheid Lt af. en lymfoïde knobbeltjes in de organen van het immuunsysteem. Bij ontstekingsprocessen en activering van immuunreacties, zowel primair als secundair (zie Immunopathologie) ,
er zijn reactieve lymfeklieren. L. t. wordt aangetast in hemoblastosen (Hemoblastosen) ,
histiocytose (Histiocytose X) x, Lymfogranulomatose ,
kwaadaardige lymfomen, paraproteïnemische hemoblastosen (paraproteïnemische hemoblastosen) .
Bibliografie: Sapin MR Immuunstructuren van het spijsverteringsstelsel, p. 123, M., 1987; hij, Principes van organisatie en patronen van structuur van de organen van het menselijke immuunsysteem. anat., hist. en embryol., t.92, nr. 2, p. 5, bibliografie.
1. Kleine medische encyclopedie. - M.: Medische Encyclopedie. 1991-96 2. Eerst gezondheidszorg. - M.: Grote Russische Encyclopedie. 1994 3. encyclopedisch woordenboek Medische termen. - M.: Sovjet Encyclopedie. - 1982-1984.
Zie wat "Lymfoïde weefsel" is in andere woordenboeken:
Lymfoïde weefsel van de wanden van de luchtwegen en het spijsverteringsstelsel- De ophoping van lymfoïde weefsel dat follikels bevat tegen de achtergrond van diffuus gelokaliseerde cellulaire elementen, die een dichtere (nodulaire) ophoping van cellen zijn, wordt amandelen (tonsillae) genoemd. Amandelen in de eerste ...... Atlas van de menselijke anatomie
- (textus, LNH) een systeem van cellen en niet-cellulaire structuren verenigd door een gemeenschappelijke functie, structuur en (of) oorsprong. Granulatieweefsel (granulationes; synoniem: granulatie, T. granulair) bindweefsel T., gevormd tijdens de genezing van weefseldefecten ... Medische Encyclopedie
- (lat. cambiumuitwisseling, verandering) de algemene naam van T., waarin intensieve celdeling plaatsvindt (bijvoorbeeld lymfoïde weefsel, darmepitheel) ... Groot medisch woordenboek
Een weefsel dat cellen produceert die beschermende reacties van het lichaam uitvoeren - lymfocyten en plasmacellen. Het is in het lichaam aanwezig in de vorm van vele discrete formaties (bijvoorbeeld lymfeklieren, amandelen, thymus en ... ... Medische termen
- (t. lymphadenoideus) zie lymfoïde weefsel ... Groot medisch woordenboek
- (t. lymphoreticularis) zie lymfoïde weefsel ... Groot medisch woordenboek
LYMFOIDDE WEEFSEL- (lymfoïde weefsel) weefsel dat cellen produceert die beschermende reacties van het lichaam uitvoeren - lymfocyten en plasmacellen. Het is in het lichaam aanwezig in de vorm van vele discrete formaties (bijvoorbeeld lymfeklieren, amandelen, thymus ... Woordenboek in de geneeskunde
- (t. lymphoideus; lymph + Grieks. eides gelijkaardig; synoniem: T. lymphadenoid, T. lymphoreticulaire) reticulaire T. met een groot aantal lymfocyten; vormt het parenchym van de lymfeklieren, milt, amandelen, thymusklier, zijn eigen plaat ... ... Groot medisch woordenboek
lymfestelsel- maakt deel uit van het cardiovasculaire systeem en vult het veneuze systeem aan, neemt deel aan de stofwisseling, reinigt cellen en weefsels. Het bestaat uit lymfebanen die de transportfuncties en organen van het immuunsysteem die de functies uitvoeren van ... ... Atlas van de menselijke anatomie
de lymfeklieren- (nodi lymphatici) de meest talrijke organen van het immuunsysteem. In het menselijk lichaam bereikt hun aantal 500. Ze bevinden zich allemaal op het pad van de lymfestroom en dragen, in afnemende mate, bij aan de verdere vooruitgang ervan. Hun belangrijkste functie is...... Atlas van de menselijke anatomie
Organen van het immuunsysteem- Het immuunsysteem zorgt voor de immuunafweer van het lichaam dankzij de cellulaire elementen van het immuunsysteem, namelijk lymfocyten en plasmacellen. Het immuunsysteem bestaat uit de lymfeklieren, milt, beenmerg, thymus of thymus... Atlas van de menselijke anatomie
Levenscyclus van immunocompetente cellen lymfoïde weefsel (B- en T-cellen en hun nakomelingen in de vorm van cellen die immunoglobulinen en antigeenherkennende en effectorlymfocyten synthetiseren) is aanzienlijk korter dan de levensduur van het organisme. In dit opzicht is de belangrijkste voorwaarde voor de implementatie van immunologische reacties de constante stroom van histogenese in het lymfoïde weefsel, waarbij de voorlopers van immunocompetente cellen prolifereren en differentiëren tot rijpe cellen.
De laatste zijn meestal zeer gespecialiseerd en heterogeen en differentiatieverschillen die verband houden met immunologische functies zijn daarin rigide vastgelegd (de klasse en specificiteit van het gesynthetiseerde immunoglobuline in B-cellen en de specificiteit van receptoren voor antigenen in T-cellen). Integendeel, onder de voorlopercellen gedurende het hele leven zijn er ook die waarvoor verschillende differentiatiemogelijkheden openstaan.
Het orgaan van immuniteit - - heeft nog een ander belangrijk kenmerk. De meeste van zijn voorlopercellen zijn mobiele, recirculerende elementen. Ze worden gevormd op plaatsen die ver verwijderd zijn van die waar ze vervolgens migreren en waar ze het nageslacht vormen van rijpe cellen.
Dus, processen, die in veel andere weefsels alleen worden uitgevoerd op het moment van leggen tijdens de morfogenese, in het lymfoïde weefsel gedurende het hele leven. Ze zorgen voor een constante verandering in de cellulaire samenstelling, een reeks gesynthetiseerde immunoglobulinen (antilichamen) en soorten immunologisch actieve lymfocyten in overeenstemming met het veranderende spectrum van antigenen die het lymfoïde weefsel binnenkomen.
op lymfoïde weefsel slaagde erin om overtuigend aan te tonen dat deze veranderingen voornamelijk of zelfs uitsluitend worden uitgevoerd door de selectie van toegewijde voorlopercellen, vooraf bepaald voor latere differentiatie, ongeacht de impact van antigenen. Bij de differentiatie van lymfoïde cellen in een volwassen organisme worden dus van nature twee stadia onderscheiden: de ontwikkeling van antigeenonafhankelijke voorlopers en de antigeenafhankelijke differentiatie van hun nakomelingen.
De eerste fase begint met een algemene hematopoietische stamcel en eindigt met toegewijde voorgangers. Het is echter niet bekend in welke volgorde de scheiding in T- en B-precursoren in dit stadium plaatsvindt, in precursors die verschillen in immunologische specificiteit en klassen van gesynthetiseerde immunoglobulinen, evenals in behorend tot verschillende subpopulaties van T-cellen. De resultaten die werden verkregen met het gebruik van transplantatie van beenmergcellen die waren gemarkeerd met chromosomale herschikkingen na bestraling, toonden met name aan dat het beenmerg stamcellen bevat die in staat zijn zichzelf in stand te houden, maar een beperkte differentiatiekracht hebben, bijvoorbeeld cellen die alleen differentiëren in myeloïde of alleen in T-cellen.
Het is echter niet gevonden terwijl stamcellen, verstoken van potentie voor myeloïde differentiatie, maar behield het vermogen om voorlopers te zijn voor zowel T- als B-cellen. Er kan daarom worden aangenomen dat zelfvoorzienende voorlopers van T-cellen (prethymische stamcellen) direct zijn afgeleid van pluripotente hematopoëtische stamcellen zonder het voorloperstadium te doorlopen dat alle lymfoïde cellen gemeen hebben. Wat betreft B-cellen, tussen hen en hematopoëtische stamcellen zijn er mogelijk helemaal geen voorlopers die in staat zijn zichzelf in stand te houden.
De kwestie van de mechanismen die werken onder opleiding van toegewijde voorlopers en het reguleren van de expressie van genen die verantwoordelijk zijn voor immunologische functies daarin, blijft open. Er zijn goede redenen om aan te nemen dat deze belangrijke fase in de ontwikkeling van immunocompetente cellen antigeenonafhankelijk is. Integendeel, de daaropvolgende ontwikkeling van reeds geëngageerde cellen en hun nakomelingen is duidelijk antigeenafhankelijk. Het zorgt voor een verandering in de cellulaire samenstelling van het lymfoïde weefsel door voornamelijk die voorlopers te vermenigvuldigen en te differentiëren waarvan de oppervlaktereceptoren de antigenen die er op dit moment in aanwezig zijn, herkennen.
Een ander mechanisme voor de selectieve proliferatie van toegewijde voorgangers geassocieerd met cellulaire interacties waarbij een door antigeen geactiveerde cel aangrenzende progenitorcellen stimuleert. In beide gevallen werken antigenen als initiatoren van veranderingen in de samenstelling van celpopulaties die het lymfoïde weefsel bewonen, en als inductoren van eiwitten die door dit weefsel worden gesynthetiseerd.
Lymfoïde weefsel (synoniem met lymfatisch weefsel) - structuren waarin de vorming van lymfocyten plaatsvindt. Deze omvatten lymfeklieren, milt, thymusklier, amandelen In het slijmvlies van het spijsverteringskanaal wordt lymfoïde weefsel weergegeven door enkele en gegroepeerde (Peyer's patches) lymfatische follikels.. Plots van lymfoïde weefsel bevinden zich in het slijmvlies van sommige organen (bronchiën, urinewegen, nieren).
Een van de belangrijkste functies van de lymfoïde organen is hun deelname aan de processen van hematopoëse (lymfopoëse). Lymfocyten kunnen, amoeboidaal bewegend, door de vaatwand gaan en fagocyteren, evenals aanleiding geven tot verschillende types cellulaire vormen, waaronder plasmacellen die betrokken zijn bij de productie van antilichamen. Een belangrijke functie van lymfoïde weefsel is geassocieerd met dit vermogen van lymfocyten - zijn deelname aan de afweerreacties van het lichaam.
de lymfeklieren - LU in de embryonale periode worden aan het einde van 2 maanden van het mesenchym langs de lymfevaten gelegd. Uit het mesenchym wordt een stroma (capsule en trabeculae-septa) gevormd en de basis van het orgaan is het reticulaire weefsel. Hematopoëtische cellen van de BMC en thymus worden al snel bevolkt in het beginnende reticulaire weefsel.
Structuur- Het orgel is boonvormig. Vanaf de convexe zijde komen de afferente lymfevaten het orgel binnen, vanaf de concave zijde verlaten de poorten de aderen die de lymfevaten uitvoeren en komen de slagaders en zenuwen binnen. Lymfeklieren zijn samengesteld uit stroma en parenchym. Stroma Het wordt weergegeven door een capsule van dichte, ongevormde SD en trabeculae-septa van losse SD die zich uitstrekken vanaf de capsule. De basis van het parenchym bestaat uit reticulair weefsel, gepenetreerd door sinussen van het bloed, en het dragen van lymfocyten op zijn lussen. Ophopingen van lymfocyten in de corticale laag (perifere zone, onder de capsule) vormen lymfatische follikels (of knobbeltjes) en in de medulla vormen pulpachtige strengen. Het lymfoïde weefsel tussen de lymfeklieren en de pulpakoorden wordt de paracorticale zone genoemd. . In de lymfatische knobbeltjes wordt een reactief centrum (of reproductiecentrum), een mantelzone onderscheiden. T-lymfocyten (40-70% van alle lymfocyten van het orgaan) bevinden zich voornamelijk in de paracotrische zone en B-lymfocyten (20-30%) - in de lymfeklieren en in de pulpakoorden.
De lymfeklieren hebben bloed sinussen:
1. Marginale sinus - tussen de capsule en de lymfeklieren.
2. De marginale sinussen lopen door in de tussenliggende of rond-nodulaire sinussen - tussen de trabecula en de lymfeklier.
3. De tussenliggende sinussen gaan verder in de cerebrale sinussen - tussen de pulpachtige koorden.
4. De cerebrale sinussen bij de poorten worden verzameld in de centrale sinus, van waaruit de lymfe wordt afgevoerd door de efferente lymfevaten.
De wand van de sinussen is bekleed met platte veelhoekige cellen, die weinig verschillen van normaal endotheel. De bekleding van de sinussen is niet continu, er blijven openingen tussen de cellen - fenestrae, het basaalmembraan is afwezig; dit alles vergemakkelijkt het binnendringen van lymfocyten in de lymfe die er doorheen stroomt.
Functies van de lymfeklieren:
1. Deelname aan lymfocytopoëse - in het lymfoïde weefsel van het orgaan worden rijpe lymfocyten en plasmocyten gevormd uit T- en B-precursoren.
2. Filtratie en zuivering van stromende lymfe.
3. Verrijking van de stromende lymfe met lymfocyten.
Morfologische verschillen van lymfeklieren bij pasgeborenen:
De capsule is dun, er zijn geen trabeculae;
Het lymfoïde weefsel is diffuus, er zijn geen duidelijke knobbeltjes en strengen;
Sinussen zijn niet gedefinieerd.
3. Testikel: bronnen van ontwikkeling, structuur, functies, regulering. Hematotesticulaire barrière.
Buiten is de testis bedekt met zijn eigen vaginale membraan, een visceraal vel van het peritoneum. Vervolgens ligt een dicht bindweefsel (eiwit) membraan. De diepe lagen van de albuginea zijn opgebouwd uit los bindweefsel dat rijk is aan bloedvaten, wat reden geeft om een ander membraan te isoleren - het vaatstelsel.
Bindweefsel septa - septa - die de testis in lobben verdeelt, strekken zich uit van de albuginea tot in de diepten van het orgel. In een van de zones van de testis verdikt de albuginea en vormt het mediastinum, waarin het netwerk van de testis zich bevindt. Het membraan van de testis, septa en mediastinum, dat wil zeggen het bindweefselgedeelte van het orgaan, vormen het stroma. Het parenchym wordt vertegenwoordigd door seminiferous ingewikkelde tubuli, daartussen bevindt zich interstitieel weefsel.
De testislobule bestaat uit takken van de tubulus seminiferi. De gekronkelde tubuli worden recht. Ze komen het mediastinum binnen en vormen hier het netwerk van de testis. Buiten is de tubulus seminiferi omgeven door een bindweefselomhulsel dat fibrocyten, collageen en elastische vezels bevat. Het basaalmembraan ligt dieper. Het bevat cellen van het spermatogeen epitheel en ondersteunende cellen (Sertoli-cellen).
Met betrekking tot het spermatogene epitheel vervullen ondersteunende cellen een trofische functie. Ze zijn langwerpig met grote, slechte chromatine, een driehoekige of ovale kern met een duidelijk zichtbare nucleolus. De cellen hebben een brede basis en een vernauwd apicaal deel dat in de tubulus is gericht. Ze hebben goed ontwikkelde agranulair endoplasmatisch reticulum, ribosomen, mitochondriën; overvloed aan insluitsels van lipiden en koolhydraten. Bevat microfilamenten en microtubuli. Ondersteunende cellen ontwikkelen zich niet tot geslachtscellen.
Het spermatogene epitheel van de tubulus seminiferi bevindt zich in verschillende stadia van differentiatie. Snermatogonia liggen in de buurt van het basaalmembraan - kleine cellen met een intens gekleurde kern gevuld met gecondenseerd chromatine.
De volgende 1-2 rijen bestaan uit primaire spermatocyten. Ze onderscheiden zich door een grote ronde kern met een goed gedefinieerd kernmembraan en een glomerulus van spiraalvormige chromosomen. De periode van groei vervangt de periode van rijping: elke primaire spermatocyt maakt twee delingen die na elkaar volgen. Na de eerste deling worden twee secundaire spermatocyten gevormd, als gevolg van de tweede deling worden twee spermatiden gevormd. Tijdens het rijpingsproces wordt het aantal chromosomen in de cellen gehalveerd en worden ze haploïde. Spermatiden zijn de kleinste cellen van het spermatogene epitheel met een kleine, zeer lichte kern en een uitgesproken nucleolus. Ze zijn meestal gerangschikt in meerdere rijen. In het lumen van de tubulus seminiferi bevinden zich cellen in het stadium van vorming en gevormd sperma.
De ruimtes tussen de tubuli zijn opgevuld met los bindweefsel. Onder de cellen en dunne vezels bevinden zich grote interstitiële endocrinocyten, die meestal in groepen voorkomen. Ze hebben een grote ronde kern met een nucleolus en een cytoplasma dat lipiden, vetten en pigmenten bevat. Deze cellen synthetiseren het hormoon testosteron. De gekronkelde tubuli worden recht. Hun wand van binnenuit is bedekt met één laag zuilvormige cellen, die qua eigenschappen vergelijkbaar zijn met de ondersteunende cellen van de ingewikkelde tubuli van de testis. Rechte buisjes zijn het begin van de uitscheidingsroutes van het zaad. Rechte buisjes komen het mediastinum binnen, waar ze een netwerk van testikels vormen. 10-30 sterk gekronkelde efferente kanalen vertrekken van dit netwerk en vormen de kop van de bijbal.